Граница перекрестка: Определение границы перекрестка
Границы перекрестка \ Акты, образцы, формы, договоры \ Консультант Плюс
]]>Подборка наиболее важных документов по запросу Границы перекрестка (нормативно–правовые акты, формы, статьи, консультации экспертов и многое другое).
Судебная практика: Границы перекрестка Открыть документ в вашей системе КонсультантПлюс:Подборка судебных решений за 2018 год: Статья 12.12 «Проезд на запрещающий сигнал светофора или на запрещающий жест регулировщика» КоАП РФ
(В.Н. Трофимов)Водитель полагал, что он был необоснованно привлечен к административной ответственности по ч. 1 ст. 12.12 КоАП РФ. При этом водитель ссылался на то, что на запрещающий сигнал светофора границу перекрестка не пересекал, остановился за стоп-линией, продолжив движение вновь на разрешающий сигнал светофора. Суд счел недоказанной вину водителя, при этом разъяснил, что на представленных материалах, в том числе обзорном фотоснимке в момент включения запрещающего сигнала светофора, последующих обзорных фотоснимках, а также видеозаписи, момент пересечения транспортным средством стоп-линии не был зафиксирован.
Подборка судебных решений за 2018 год: Статья 56.10 «Заключение соглашения об изъятии недвижимости для государственных или муниципальных нужд» ЗК РФ
(ООО юридическая фирма «ЮРИНФОРМ ВМ»)Принимая решение о принудительном изъятии объектов недвижимости для муниципальных нужд с установлением размера компенсации убытков, причиненных изъятием, суд в порядке пункта 10 статьи 56.10 ЗК РФ, пункта 28 Постановления Пленума ВАС РФ от 24.03.2005 N 11 установил, что предусмотренная процедура для изъятия для муниципальных нужд земельных участков и расположенных на них объектов недвижимости истцом соблюдена надлежащим образом, спорные объекты подлежат изъятию из собственности ответчика, поскольку находятся в границах проекта транспортно-пересадочного узла, которым учтен проект планировки и межевания территории транспортно-разворотной площадки, перекрестка, а исключение из списка изымаемых объектов объекта культурного наследия согласовано с уполномоченным органом как не подлежащий сносу, при этом на основании заключения судебной экспертизы определена общая стоимость возмещения убытков ответчику в связи с изъятием объектов недвижимости, включая рыночную стоимость права аренды земельного участка.
 Статьи, комментарии, ответы на вопросы: Границы перекрестка Открыть документ в вашей системе КонсультантПлюс:
Статьи, комментарии, ответы на вопросы: Границы перекрестка Открыть документ в вашей системе КонсультантПлюс:Статья: Конкуренция на финансовом рынке
(«Конкурентное право», 2018, N 4)Разделение границы между перекрестными продажами и навязыванием услуг достаточно условное, поэтому финансовые участники должны находить баланс между этим. Было бы также правильным использовать возможность, чтобы не все услуги в рамках перекрестных услуг были платные, платным должен быть именно комплекс оказываемых услуг. То есть на вышеуказанном примере, если клиент соглашается взять кредит, то банк предоставит клиенту бесплатное обслуживание его счета.Нормативные акты: Границы перекрестка
определение ПДД. Разобрано очень подробно!
Что такое перекресток? Давайте сначала посмотрим это определение в ПДД.
“Перекресток” – место пересечения, примыкания или разветвления дорог на одном уровне, ограниченное воображаемыми линиями, соединяющими соответственно противоположные, наиболее удаленные от центра перекрестка начала закруглений проезжих частей. Не считаются перекрестками выезды с прилегающих территорий.
На фото внизу – обычный перекресток.
А теперь разберем это определение более подробно.
Самое важное, что нужно запомнить, что перекресток – пересечение примыкание дорог на одном уровне.
Ключевая фраза – пересечение на одном уровне!
Могут быть пересечения на разных уровнях, скажем эстакада, но это не перекресток.
Так с перекрестком разобрались, давайте теперь посмотрим какие виды перекрестков бывают.
Регулируемый перекресток – определение ПДД.
Перекресток, где очередность движения определяется сигналами светофора или регулировщика, считается регулируемым.
То есть если на перекрестке есть светофор или регулировщик, то этот перекресток – регулируемый.
Или регулировщик.
Нерегулируемый перекресток определение ПДД.
Если нет ни того ни другого, то считаем этот перекресток нерегулируемым и будем руководствоваться знаками приоритета (если они есть), а если нет, то правилом помехи справа.
Так же нерегулируемым считается перекресток, если светофор на нем перешел в режим желтого моргающего сигнала.
Границы перекрестка.
Нужно знать и уметь находить границы перекрестка.
Для этого сначала найдем максимально удаленные точки закруглений проезжих частей.
границы перекрестка.Не нужно путать границы перекрестка и границы пересечений проезжих частей.
Для того что бы найти пересечение проезжих частей на перекрестке, нужно всего лишь продлить каждую проезжую часть.
Пересечение проезжих частей.На этом с перекрестком все, подписывайтесь на нашу группу ВК, там найдете много интересного.
Самое частое заблуждение водителей в правилах проезда перекрестка
Масса аварий, нарушений ПДД и претензий водителей друг к другу из-за этого недопонимания
Активно, что уж греха таить, штрафуя водителей за нарушения правил проезда перекрестков, мы столкнулись с явным пробелом в знаниях в такой, казалось бы, стандартной области правил дорожного движения.
Нет смысла приводить выдержки из ПДД, все вы этот документ хорошо знаете. Но, если не поняли данные постулаты, то попробуем объяснить своими словами. Извините, конечно, что самонадеянно обвиняем почти всю страну в непонимании, этот момент у вас уже, наверно, вызвал раздражение. Но через нас проходят те «счастливые бумажки», по которым вы пишите жалобы на неправильную оценку ваших действий на перекрестке. Через нас проходит и разбор ваших полетов, когда водители не уступают друг другу дорогу на перекрестке: один, видя, что едет на зеленый, другой – потому что законно заканчивает маневр на красный.
Даже на телевидении часто приходится слышать оговорки по Фрейду. Например, в этом выпуске «Утро России» на минуте 08.19 диктор говорит что-то типа: Наконец, Управление ГИБДД разъяснило, что является выездом на перекресток – пересечение бампером стоп-линии.
Итак, выезд за стоп-линию – это всего лишь выезд за стоп-линию — физически, и никакого отношения к выезду или пересечению перекрестка он не имеет. Если вы пересекли бампером стоп-линию на запрещающий сигнал светофора, то получите штраф именно по этой статье. А, если выехали на разрешающий и не смогли продолжить движение, то стойте и не дергайтесь, потому что никаких дополнительных прав этот момент вам не дает. И совсем другое дело с перекрестком.
У него есть еще одна очень важная, причем, критически важная незримая линия. К сожалению, ее не отмечают на асфальте, но она будет поважнее стоп-линии – это граница самого перекрестка. И она чаще не совпадает со стоп-линией.
За пересечение границы перекрестка на запрещающий сигнал светофора вы попадаете на более вескую статью. Но важнее знать, какие ОГРОМНЫЕ ПРАВА дает пересечение этой линии на разрешающий сигнал светофора. В этом случае, даже в случае, если задержался в начале своего пути и даже после появления запрещающего сигнала, водитель ТС имеет право и должен закончить свой маневр – пересечь перекресток. Разница непонятна? Если вы спокойно трогаетесь на зеленый и вдруг встречаетесь с автомобилем, летящим вам наперекор, то есть высокая вероятность, что нарушаете вы!
В этом случае, даже в случае, если задержался в начале своего пути и даже после появления запрещающего сигнала, водитель ТС имеет право и должен закончить свой маневр – пересечь перекресток. Разница непонятна? Если вы спокойно трогаетесь на зеленый и вдруг встречаетесь с автомобилем, летящим вам наперекор, то есть высокая вероятность, что нарушаете вы!
Причем, определить эту вероятность очень сложно, потому что никто не знает (если нет камер), от какой точки стартанула вошедшая в вас машина: от стоп-линии или от границы перекрестка? Т.е., скорее всего, виновного не установят. Но будет ли вам от этого легче?
Поэтому мы, имея огромную негативную статистику по таким делам, настоятельно рекомендуем:
Не сильно торопиться трогаться при появлении разрешающего сигнала светофора.
Вопреки правилам, по возможности не пересекать перекресток на «запрещающий», если вы уже выехали за его границу. Просто потому что первой рекомендацией многие пренебрегают.

Источник: https://goal.ru/security-systems-video/samoe-chastoe-zabluzhdenie-voditeley/
Данный материал является частной записью члена сообщества Club.CNews.
Редакция CNews не несет ответственности за его содержание.
Экспертная оценка термина «перекресток» Текст научной статьи по специальности «Право»
13. Arhangel’skij, V. M. Avtomobil’nye dvigateli / V. M. Arhangel’skij, M. M. Vihert, A. N. Voinov, YU. A. Stepanov. M.:Mashinostroenie, 1967. 496 s.
14. Handov, Z. A. Sudovye dvigateli vnutrennego sgoraniya (teoriya) / Z. A. Handov. M. Transport, 1969. 304 s.
15. Mamedova, M. D. Rabota dizelya na szhiz-hennom gaze / M. E. Mamedova. M.: Mashinostroenie, 1980.149 s.
16. Vedruchenko, V. R. Metodika indicirovaniya sredneoborotnogo sudovogo dizelya / V. R. Vedruchenko // Peredovoj opyt i novaya tekhnika. CB NTI MRF. Vyp. 4. M., 1981. S. 37 — 41.
Ведрученко Виктор Родионович (Омск, Россия) — доктор технических наук, профессор, профессор кафедры Теплоэнергетика ФГБОУ ВО ОмГУПС (644046, г.Омск, пр. Маркса, 35, e-mail: [email protected]).
Крайнов Василий Васильевич (Омск, Россия) -кандидат технических наук, доцент кафедры Теплоэнергетика ОмГУПС (644046, г.Омск, пр. Маркса, 35, e-mail: [email protected]).
Лазарев Евгений Сергеевич (Омск, Россия) -преподаватель кафедры Теплоэнергетика ФГБОУ ВО ОмГУПС (644046, г.Омск, пр. Маркса, 35, e-mail: [email protected]).
УДК 656.05
Литвинов Павел Васильевич (Омск, Россия) -аспирант очной формы обучения кафедры Теплоэнергетика ФГБОУ ВО ОмГУПС (644046, г. Омск, пр.Маркса, 35, e-mail: [email protected]).
Омск, пр.Маркса, 35, e-mail: [email protected]).
Victor R. Vedruchenko (Omsk, Russian Federation) — Doctor of Technical Sciences, Professor, Department of Heat Energy, Omsk State Transport University (644046, Marks avenue, 35, Omsk, e-mail: [email protected]).
Vasily V. Krainov (Omsk, Russian federation) -, Candidate of Technical Sciences, Associate Professor, Department Of Heat Energy, Omsk State Transport University (644046, Marks avenue, 35, Omsk, email: [email protected]).
Yevgeny S. Lazarev (Omsk, Russian Federation) — Lector, Department of Heat Energy, Omsk State Transport University (644046, Marks avenue, 35, Omsk, e-mail [email protected]).
Pavel V. Litvinov (Omsk, Russian Federation) -post graduate student of the Heat Energy Department of the Omsk State Transport University (644046, Marks avenue, 35, Omsk, e-mail: p_vasilich55@mail. ru).
ru).
ЭКСПЕРТНАЯ ОЦЕНКА ТЕРМИНА «ПЕРЕКРЕСТОК»
В.А. Городокин, З.В. Альметова, В. Д. Шепелев ФГАОУ ВО «ЮУрГУ (НИУ)», г. Челябинск, Россия
Аннотация. Статистические данные ГИБДД свидетельствуют о том, что самыми распространенными видами дорожно-транспортных происшествий стали столкновения транспортных средств, большинство из которых происходят на перекрестках дорог. Объективное расследование обстоятельств дорожно-транспортных происшествий и установление истинного виновника происшествия является основной задачей органов и лиц, проводящих исследование обстоятельств ДТП. Вместе с тем, проведение исследования обстоятельств происшествия неразрывно связано с однозначным пониманием того, где, в каком месте произошло дорожно-транспортное происшествие и кто из его участников пользовался преимущественным правом на движение. Решение поставленных вопросов, зачастую, затруднено в связи с недостаточно точными формулировками терминов, используемых в Правилах дорожного движения Российской Федерации. В частности этот недостаток касается термина «перекресток». В различной литературе, посвященной вопросам безопасности движения, приводятся примеры, позволяющие определить конфигурацию и границы перекрестка. Однако, подавляющее большинство приведенных примеров базируется на наиболее простой форме трех- или четырех стороннего перекрестка. В данной работе авторами проведен детальный анализ и сделан вывод о необходимости изменения существующего термина «перекресток».
В частности этот недостаток касается термина «перекресток». В различной литературе, посвященной вопросам безопасности движения, приводятся примеры, позволяющие определить конфигурацию и границы перекрестка. Однако, подавляющее большинство приведенных примеров базируется на наиболее простой форме трех- или четырех стороннего перекрестка. В данной работе авторами проведен детальный анализ и сделан вывод о необходимости изменения существующего термина «перекресток».
Ключевые слова: дорожно-транспортное происшествие, перекресток, безопасность движения, правила дорожного движения, границы перекрестка.
Введение
Согласно статистическим данным ГИБДД, по итогам 2015 года снизились все основные показатели аварийности. Число погибших в дорожно-транспортных происшествиях
уменьшилось на 14,7 %. Тем не менее, всего в 2015 году на российских дорогах погибли 23 114 человек. Общее количество ДТП уменьшилось на 8,2%, всего было зарегистрировано 184 тыс. ДТП. При этом, число раненых сократилось на 8,6%, а ранения на дорогах получили 231 197 человек. По вине водителей произошло 157 943 ДТП (-9,2%), в которых ранено 207 985 человек (-9,2%), погибло 19 011 человек (-16,9%). Самыми распространенными видами дорожно-транспортных происшествий стали столкновения транспортных средств (75 266 случаев), наезд на пешехода (56 918) и наезд на препятствие (11 842) [1].
Тем не менее, всего в 2015 году на российских дорогах погибли 23 114 человек. Общее количество ДТП уменьшилось на 8,2%, всего было зарегистрировано 184 тыс. ДТП. При этом, число раненых сократилось на 8,6%, а ранения на дорогах получили 231 197 человек. По вине водителей произошло 157 943 ДТП (-9,2%), в которых ранено 207 985 человек (-9,2%), погибло 19 011 человек (-16,9%). Самыми распространенными видами дорожно-транспортных происшествий стали столкновения транспортных средств (75 266 случаев), наезд на пешехода (56 918) и наезд на препятствие (11 842) [1].
Наиболее часто дорожно-транспортные происшествия (ДТП) происходят на узлах улично-дорожной сети, к которым в первую очередь относятся перекрестки дорог. Не вызывает сомнения тот факт, что объективное расследование обстоятельств ДТП и установление истинного виновника происшествия является основной задачей органов и лиц, проводящих исследование обстоятельств ДТП. Проведение исследования обстоятельств происшествия неразрывно связано с однозначным пониманием того, где, в каком месте ДТП и кто из его участников пользовался преимущественным правом на движение. Однако, решение поставленных вопросов, зачастую, затруднено в связи с недостаточно точными формулировками терминов, используемых в Правилах дорожного движения Российской Федерации (ПДД РФ).
Однако, решение поставленных вопросов, зачастую, затруднено в связи с недостаточно точными формулировками терминов, используемых в Правилах дорожного движения Российской Федерации (ПДД РФ).
Определение границ перекрестка
Так, согласно принятой в Российской Федерации терминологии, «Перекресток» — место пересечения, примыкания или разветвления дорог на одном уровне, ограниченное воображаемыми линиями, соединяющими соответственно противоположные, наиболее удаленные от центра перекрестка начала закруглений проезжих частей. Не считаются перекрестками выезды с прилегающих территорий».
Как следует из приведенного выше термина, под перекрестком понимается достаточно обширная территория, образованная пересекающимися, разветвляющимися или примыкающими проезжими частями. По сути, это та часть дороги, которая ограничена прямыми линиями, проходящими в зоне окончания и начала закруглений проезжих частей.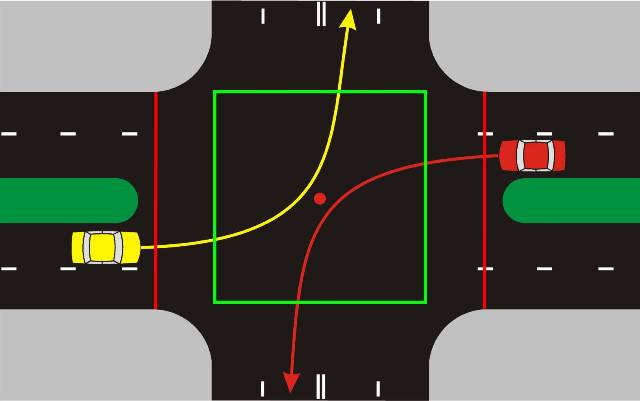
Вместе с тем, в практической деятельности в первую очередь требуется устанавливать не сами проезжие части и факт их пересечения, примыкания и разветвления, а границы этих форм узлов автомобильных дорог. К сожалению, в действующей редакции ПДД, равно, как во всех предыдущих редакциях, разъяснению термина «граница пересечения проезжих частей» или «граница пересекаемой проезжей части» не дано. При разборе указанных выше терминов следует указать на несовпадение существующих теоретических формулировок и классификации, данной в специальной технической литературе, посвященной дорожному проектированию, терминологии, используемой в ПДД. Так, согласно принятой в технической литературе классификации, «автомобильные дороги, образующие дорожную сеть страны, могут пересекаться между собой, примыкать друг к другу или разветвляться на два или несколько направлений» [2].
Отличие терминологии, используемой в ПДД, от применяемой в технической литературе состоит в том, что как пересечение, так и примыкание или разветвление, представляющих собой узел автомобильных дорог, именуются одним термином «пересечение» проезжих частей. С технической точки зрения, различают узлы автомобильных дорог в одном или в разных уровнях (двух, трех, четырех и более). Узлы, состоящие из автомобильных дорог, пролегающих в разных уровнях, называются «транспортными развязками» и, соответственно, «перекрестками», с точки зрения ПДД, не являются. В свою очередь, применительно к терминологии ПДД, узел автомобильных дорог, пролегающих в одном уровне, подпадает под термин «перекресток» [3].
С технической точки зрения, различают узлы автомобильных дорог в одном или в разных уровнях (двух, трех, четырех и более). Узлы, состоящие из автомобильных дорог, пролегающих в разных уровнях, называются «транспортными развязками» и, соответственно, «перекрестками», с точки зрения ПДД, не являются. В свою очередь, применительно к терминологии ПДД, узел автомобильных дорог, пролегающих в одном уровне, подпадает под термин «перекресток» [3].
В ПДД термин «перекресток» или производные этого слова употребляется 68 раз и, как это следует из приведенной выше классификации, «перекресток» является более широким понятием и, соответственно, охватывает такой узел, как «пересечение проезжих частей». Вывод о том, что в ПДД все узлы, представляющие собой пересечение, примыкание или разветвление, именуются одним термином «пересечение» проезжих частей, подтверждается формулировками различных требований ПДД, например, фрагментом п.12.4, согласно которому, «Остановка запрещается: . ..на пересечении проезжих частей и ближе 5 м от края пересекаемой проезжей части, за исключением стороны напротив бокового проезда трехсторонних пересечений (перекрестков), имеющих
..на пересечении проезжих частей и ближе 5 м от края пересекаемой проезжей части, за исключением стороны напротив бокового проезда трехсторонних пересечений (перекрестков), имеющих
сплошную линию разметки или разделительную полосу…». Как видно из приведенного примера, речь идет об узле автомобильных дорог, представляющих собой или примыкание или разветвление, однако, как один, так и другой вариант названы «пересечением».
Вместе с тем, как указано выше, с точки зрения выполнения требований ПДД, наибольший интерес представляет не сам объект — пересечение проезжих частей, а его границы. В частности, определение границы проезжей части, на которую осуществляется въезд при пересечении, потребуется для решения вопроса о соответствии или несоответствий действий водителя транспортного средства требованиям п.6.13 ПДД. Иными словами, ПДД требуют от водителя в первую очередь определить ближайшую границу перекрестка, затем соотнести с ней нанесенную стоп-линию, и, при отсутствии стоп-линии, следующим шагом определить ближайшую границу проезжей части, которую он намерен пересечь, или на которую выехать.
Дальнюю границу пересечения проезжих частей водитель должен установить с целью выполнения требований ч.1 п.8.6 ПДД, согласно которым «Поворот должен осуществляться таким образом, чтобы при выезде с пересечения проезжих частей транспортное средство не оказалось на стороне встречного движения».
В связи с вышеизложенным, возникает несколько вопросов:
1.Как водитель должен устанавливать границы пересечения проезжих частей?
2.Что является границей пересечения проезжих частей?
И, если, как было указано выше, в ПДД РФ существует термин «перекресток» и дается его понимание, то, что такое пересечение проезжих частей и, что является границей пересечения проезжих частей, в нормативно-правовом акте нет. При этом термин «перекресток» представляет собой несколько более развернутое понятие, чем то, которое дано в Конвекции о дорожном движении: «Intersection» means any level crossroad, junction or fork, including the open areas formed by such crossroads, junctions or forks», что означает «Перекресток» — любое пересечение, примыкание или разветвление дорог в одном уровне, включая территорию, образуемую такими пересечениями, примыканиями или разветвлениями» (перевод официального текста скорректирован авторами) [4]. Нельзя сказать, что приведенная формулировка в международном правовом документе в полной мере позволяет определить границы пере-
Нельзя сказать, что приведенная формулировка в международном правовом документе в полной мере позволяет определить границы пере-
крестка. Во всяком случае, непонятно о какой территории и ее протяженности идет речь, которая охватывается термином и образована в результате пересечений, примыканий и разветвлений дорог.
Таким образом, не вызывает сомнения тот факт, что существующий термин «перекресток» должен быть изменен. Еще одним доводом в пользу того, что существующая формулировка не удовлетворяет современным конструктивным особенностям перекрестков, является внедрение на перекрестках направляющих островков и специальных полос, выполненных в обход светофорных объектов. Наличие указанных полос, места съезда на них и места слияния с основным потоком, выходят далеко за пределы начала или окончания закруглений проезжих частей. Сложная конфигурация перекрестка ведет к тому, что определение его границы или представляет значительную сложность, или технически невозможно [5].
Вместе с тем, принимая во внимание тот факт, что границы пересечения проезжих частей важны, однако, не могут выходить за границы перекрестка, а само пересечение проезжих частей является его составной частью, рассмотрим вопрос о том, имеет ли принципиальное значение определение границы перекрестка.
Рассмотрим, в каких случаях применяется термин «Перекресток»,
• при указании участникам движения на ориентир при переходе дороги;
• при указании на форму организации движения в зоне пересечения проезжих частей;
• для соотнесения места установки дорожных знаков и границы перекрестка;
• при определении места запрещения определенных действий;
• при регламентации определенных действий, в зоне пересечения проезжих частей
• при нанесении разметки в границах перекрестка;
• при ограничении зоны действия дорожных знаков;
• при указании на маршрут движения, в случае запрещения определенных действий на перекрестке.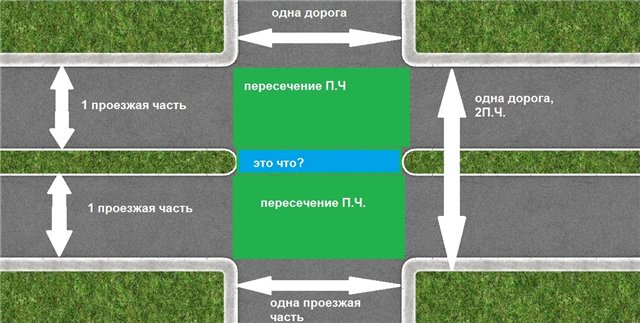
При этом, нередко в ПДД одновременно применяются оба термина, например в п.13.2. «Запрещается выезжать на перекресток или пересечение проезжих частей, если образовался затор, который вынудит водителя остановиться, создав препятствие для движения транспортных средств в поперечном на-
правлении». Вместе с тем, в приведенной формулировке достаточно употребления одного термина — «пересечение проезжих частей». Выезд (правильнее говорить «въезд», т.к. согласно правилам русского языка, выехать можно откуда-то, а куда-то можно въехать) на перекресток в подавляющем числе случаев не препятствует движению транспортного потока в конфликтующем направлении. Помеха в данном случае может быть создана пешеходам, но это в основном будет зависеть от расположения пешеходного перехода относительно границ пересекаемой проезжей части и является темой отдельного обсуждения [6].
В другом случае формальная граница перекрестка не имеет какого-либо технического или правового значения, например, п. 13.3. «Перекресток, где очередность движения определяется сигналами светофора или регулировщика, считается регулируемым. При желтом мигающем сигнале, неработающих светофорах или отсутствии регулировщика перекресток считается нерегулируемым, и водители обязаны руководствоваться правилами проезда нерегулируемых перекрестков и установленными на перекрестке знаками приоритета».
13.3. «Перекресток, где очередность движения определяется сигналами светофора или регулировщика, считается регулируемым. При желтом мигающем сигнале, неработающих светофорах или отсутствии регулировщика перекресток считается нерегулируемым, и водители обязаны руководствоваться правилами проезда нерегулируемых перекрестков и установленными на перекрестке знаками приоритета».
Еще больший вопрос вызывает требование п.13.7 ПДД, согласно которому «Водитель, въехавший на перекресток при разрешающем сигнале светофора, должен выехать в намеченном направлении независимо от сигналов светофора на выходе с перекрестка. Однако, если на перекрестке перед светофорами, расположенными на пути следования водителя, имеются стоп-линии (знаки 6.16), водитель обязан руководствоваться сигналами каждого светофора». Проблема заключается в том, что при достаточно сложной конфигурации перекрестка и значительной его протяженности, не исключен факт въезда в границы перекрестка на разрешающий сигнал светофора и достижение границ пересекаемой проезжей части (при высокой загрузке перекрестка) на запрещающий сигнал. С точки зрения здравого смысла и безопасности дорожного движения, наиболее целесообразно водителю остановить транспортное средство (если такая техническая возможность у него имеется) до границы пересекаемой проезжей части, а не осуществлять въезд на пересечение при запрещающем сигнале [7]. Однако, вопреки здравому смыслу, согласно действующим требованиям ПДД, водитель должен продолжить движение, чем, вероятнее всего, создаст и помеху, и опасность водителям транспортных
С точки зрения здравого смысла и безопасности дорожного движения, наиболее целесообразно водителю остановить транспортное средство (если такая техническая возможность у него имеется) до границы пересекаемой проезжей части, а не осуществлять въезд на пересечение при запрещающем сигнале [7]. Однако, вопреки здравому смыслу, согласно действующим требованиям ПДД, водитель должен продолжить движение, чем, вероятнее всего, создаст и помеху, и опасность водителям транспортных
средств, начинающим движение на разрешающий сигнал светофора.
Не меньший вопрос вызывают и требования п.13.9 ПДД, согласно которым «На перекрестке неравнозначных дорог водитель транспортного средства, движущегося по второстепенной дороге, должен уступить дорогу транспортным средствам, приближающимся по главной, независимо от направления их дальнейшего движения». В данном случае проблема состоит в том, что разъяснение термина «второстепенная дорога», практически, отсутствует. Только в одном случае Правила дают возможность понять, что въезд на перекресток осуществляется с второстепенной дороги — когда примыкающая или пересекающая дорога является грунтовой, т.е. дорогой, на которой отсутствует вообще какое-либо усовершенствованное покрытие. Во всех других случаях, въезд производится просто на главную дорогу, понятие которой дано, однако ее обозначение на дороге, пересекающей или примыкающей к ней, не обозначается иным способом, как дорожными знаками 2.4 или 2.5. Отсутствие указанных знаков не позволяют водителю идентифицировать дорогу, которую он намерен пересечь, как главную, а соответственно, ту, по которой он движется, второстепенной [8].
Только в одном случае Правила дают возможность понять, что въезд на перекресток осуществляется с второстепенной дороги — когда примыкающая или пересекающая дорога является грунтовой, т.е. дорогой, на которой отсутствует вообще какое-либо усовершенствованное покрытие. Во всех других случаях, въезд производится просто на главную дорогу, понятие которой дано, однако ее обозначение на дороге, пересекающей или примыкающей к ней, не обозначается иным способом, как дорожными знаками 2.4 или 2.5. Отсутствие указанных знаков не позволяют водителю идентифицировать дорогу, которую он намерен пересечь, как главную, а соответственно, ту, по которой он движется, второстепенной [8].
Рассмотрим случаи, когда граница перекрестка имеет значение. Так, согласно требованиям п.6.13 ПДД «При запрещающем сигнале светофора (кроме реверсивного) или регулировщика водители должны остановиться перед стоп-линией (знаком 6.16), а при ее отсутствии:
• на перекрестке — перед пересекаемой проезжей частью (с учетом пункта 13. являющейся предполагаемой границей пересекаемой проезжей части. По мнению авторов работы, указанный пункт Правил должен трактоваться следующим образом: «При запрещающем сигнале
являющейся предполагаемой границей пересекаемой проезжей части. По мнению авторов работы, указанный пункт Правил должен трактоваться следующим образом: «При запрещающем сигнале
светофора (кроме реверсивного) или регулировщика водители должны остановиться перед стоп-линией (знаком 6.16), а при ее отсутствии:
• на перекрестке — водитель должен остановиться перед пересекаемой проезжей частью (с учетом пункта 13.7 Правил), не создавая помех пешеходам;
• перед железнодорожным переездом -остановиться необходимо в соответствии с пунктом 15.4 Правил;
• в других местах — останавливаться следует перед светофором или регулировщиком, не создавая помех транспортным средствам и пешеходам, движение которых разрешено».
Рис. 1. Изображение места остановки транспортных средств, согласно требованиям п. 6.13 ПДД, при запрещающем сигнале светофора и отсутствии стоп-линии: А В С Р Е Р К 1_ — границы перекрестка; М N — границы пересекаемой проезжей части; Н — группа транспортных средств
6.13 ПДД, при запрещающем сигнале светофора и отсутствии стоп-линии: А В С Р Е Р К 1_ — границы перекрестка; М N — границы пересекаемой проезжей части; Н — группа транспортных средств
При данной трактовке, учитывая тот факт, что стоп-линия отсутствует не на перекрестке (рис.1), а в другом месте, транспортные средства должны остановиться перед светофором, что, собственно, водителями указанной на схеме группы транспортных средств «Н» и сделан. Иными словами, водители указанной группы транспортных средств руководствовались не требованиями ПДД, а здравым смыслом [9].
Определенный интерес, с точки зрения определения границы перекрестка, представляют требования п.8.8 Правил, согласно которым «При повороте налево или развороте вне перекрестка водитель безрельсового транспортного средства обязан уступить дорогу встречным транспортным средствам и трамваю попутного направления. Если при развороте вне перекрестка ширина проезжей части недостаточна для выполнения маневра из крайнего левого положения, его допускается производить от правого края проезжей части (с правой обочины). При этом водитель
должен уступить дорогу попутным и встречным транспортным средствам».
Из приведенного выше текста следует, что при совершении маневра «разворот» в случаях, когда ширина проезжей части не позволяет выполнить его из крайнего левого положения, маневр допускается производить из иного положения, при этом необходимо уступать дорогу попутным и встречным транспортным средствам, не исключая трамвай. Если данное требование сравнить с требованиями п.8.7 Правил, согласно которым «Если транспортное средство из-за своих габаритов или по другим причинам не может выполнить поворот с соблюдением требований пункта 8.5 Правил, допускается отступать от них при условии обеспечения безопасности движения и если это не создаст помех другим транспортным средствам», то можно выявить ряд совпадений, позволяющих при некотором незначительном изменении и объединении формулировок признать несущественной разницу в обязанностях водителя ма-
неврирующего транспортного средства при совершения маневра «поворот» или разворот» в пределах перекрестка или вне его [10]. При данных обстоятельствах можно сделать вывод об отсутствии необходимости установления границы перекрестка.
Анализируя требования п.8.12 Правил, согласно которым «Движение транспортного средства задним ходом разрешается при условии, что этот маневр будет безопасен и не создаст помех другим участникам движения. При необходимости водитель должен прибегнуть к помощи других лиц. Движение задним ходом запрещается на перекрестках и в местах, где запрещен разворот согласно пункту 8.11 Правил», можно сделать вывод о некорректности введенного запрета. Вне всякого сомнения, двигаться через перекресток задним ходом следует запретить, но как быть в случае, если двигавшееся впереди транспортное средство остановились в пределах перекрестка не в связи с необходимостью предоставления приоритета, а по иной причине (например, неисправность, не позволяющая продолжить движение, или столкновение с другим транспортным средством). При этом дистанция до остановившегося транспортного средства не позволяет произвести его объезд. Водителю необходимо продолжать стоять или допустимо совершить маневр, связанный первоначально с движением задним ходом и последующим объездом? Видимо, следует признать допустимым второй вариант, но как быть с запретом двигаться задним ходом в пределах перекрестка? Вероятнее всего, во-первых, следует изменить формулировку п.8.12 Правил и запретить движение задним ходом не на перекрестке, а с целью пересечения проезжих частей [11].
Следующий пункт, подвергаемый анализу 9.2 ПДД «На дорогах с двусторонним движением, имеющих четыре или более полосы, запрещается выезжать для обгона или объезда на полосу, предназначенную для встречного движения. На таких дорогах повороты налево или развороты могут выполняться на перекрестках и в других местах, где это не запрещено Правилами, знаками и (или) разметкой». Более неудачную формулировку трудно представить. Написав фразу «.на перекрестках и в других местах, где это не запрещено Правилами, знаками и (или) разметкой», законодатель, по сути, допустил выполнение поворота и разворота «везде», где это не запрещено Правилами, знаками и (или) разметкой. Иными словами, на проезжих частях, имеющих ширину, позволяющую
двигаться по 4, 6 и 8 полосам в обоих направлениях, в случае, когда дорожная разметка плохо различима в силу физического износа или из-за наличия снега, грязи и т.п., водитель получает право на выполнение вне перекрестка маневров, как «поворот налево» с пересечением всех встречных полос, так и «разворот», который представляет не меньшую опасность. И стоит ли говорить о той помехе, а, зачастую, и об опасности, которую создает транспортное средство, остановившееся в крайней левой полосе на перегоне между перекрестками, на время ожидания водителем возможности совершить маневр, указанный выше. И все это допустимо, согласно действующим ПДД. При грамотной постановке требования, поворот налево или разворот на проезжих частях, указанной выше ширины, следует разрешить только на пересечении проезжих частей или в местах специально оборудованных и организованных для этой цели, и кроме этого, обозначенных соответствующим образом [12]. Таким образом, в анализируемом пункте Правил термин «перекресток» может быть или заменен на «пересечение проезжих частей», или в существующей формулировке потерял смысл и не имеет принципиального значения с точки зрения границ перекрестка.
Также теряет смысл граница перекрестка в требованиях п. 13.9. Правил, согласно которым «На перекрестке неравнозначных дорог водитель транспортного средства, движущегося по второстепенной дороге, должен уступить дорогу транспортным средствам, приближающимся по главной, независимо от направления их дальнейшего движения. На таких перекрестках трамвай имеет преимущество перед безрельсовыми транспортными средствами, движущимися в попутном или встречном направлении по равнозначной дороге, независимо от направления его движения.В случае если перед перекрестком с круговым движением установлен знак 4.3 в сочетании со знаком 2.4 или 2.5, водитель транспортного средства, находящегося на перекрестке, пользуется преимуществом перед выезжающими на такой перекресток транспортными средствами».
Используется термин «перекресток» в разделе ПДД, касающийся дорожной разметки и дорожных знаков. При этом, аналогично, не имеет значения формальная граница перекрестка при нанесении дорожной разметки 1.7 (прерывистая линия с короткими штрихами и равными им промежутками) — обозначающая полосы движения в пределах перекрестка. В данном случае, учитывая тот факт,
что перекресток образован пересекающимися проезжими частями и кривыми, соединяющими эти проезжие части, разметка 1.7 должна наноситься на всем протяжении кривой, ограничивающей, в зависимости от направления поворота, правую или левую границу стороны проезжей части.
Также теряет смысл определение границ перекрестка и имеет только ориентирующее значение наносимая разметка 1.18 — указывающая разрешенные на перекрестке направления движения по полосам.
Таким образом, анализ применяемого в ПДД РФ термина «перекресток» показывает, что применение его в подавляющем числе случаев или носит ориентирующий характер или может быть сведено к понятию термина «пересечение проезжих частей». И тот и другой вариант сводят на нет необходимость определения границы перекрестка.
Выводы
Проведенный анализ термин «перекресток» показал, что принятые и используемые в ПДД формулировки требований, предъявляемых к водителям транспортных средств, изобилуют множеством погрешностей, неточностей и просто ошибок, что позволяет в некоторых случаях двояко интерпретировать соответствующие положения. Данное положение дел недопустимо, т.к. с одной стороны вводит в заблуждение участников дорожного движения, с другой, затрудняет или исключает возможность объективного исследования обстоятельств дорожно-транспортного происшествия. На основании проведенного исследования, предлагается принять термин «перекресток» в следующей формулировке: «перекресток» — территория, образованная в одном уровне пересекающимися, примыкающими или разветвляющимися проезжими частями, а также закруглениями, соединяющими указанные проезжие части. Принятие предложенной формулировки позволит устранить неопределенность в решении множества вопросов, возникающих при расследовании обстоятельств дорожно-транспортных происшествий, имевших место в пределах перекрестка, что в свою очередь позволит принять объективное решение о виновности того или иного участника происшествия.
Библиографический список
1. ГИБДД утверждает, что все показатели аварийности снизились. — Режим доступа: http://www.zr.ru/content/news/845054-gibdd-utverzhdaet-chto-vse-pokazateli-avarijnosti-snizilis/ (Дата обращения 20.10.2016).
2. Гохман, В.А. Пересечение и примыкания автомобильных дорог : учеб. пособие для вузов / В.А. Гохман и др. — М. : Высшая школа, 1977. -310 с.
3. Правила дорожного движения Российской Федерации: по состоянию на 21.01.2015г. — М. : ООО ИДТР, 2015. — 64 с.
4. Конвенция о дорожном движении. Европейское соглашение, дополняющее Конвенцию о дорожном движении, открытую для подписания в Вене 8 ноября 1968 года (Женева 1 мая 1971 г.). -М. : Ассоциация международных автомобильных перевозчиков, 1994. — 53 с.
5. Кременец, Ю.А. Технические средства регулирования дорожного движения / Ю.А. Кременец, М.П. Печерский. — М. : Транспорт, 1981. — 252 с.
6. Городокин, В.А. О некоторых проблемах безопасности пешеходов / В.А. Городокин, З.В. Апьметова // Модернизация и научные исследования в транспортном комплексе. — 2015. — № 1. — С. 231-237.
7. Суворов, Ю.Б. Судебная дорожно-транспортная экспертиза. Судебно-экспертная оценка действий водителей и других лиц, ответственных за обеспечение безопасности дорожного движения, на участках ДТП / Ю.Б. Суворов. — М. : Экзамен, 2003. — 208 с.
8. Городокин, В.А. Правовые, технические и виктимологические проблемы обеспечения безопасности на нерегулируемых пешеходных переходах / В.А. Городокин, А.В Майоров. — Челябинск : Цицеро, 2012. — 130 с.
9. Селиванов, Н.А. Расследование дорожно-транспортных происшествий / Н.А. Селиванов,
A.И. Дворкин, Б.Д. Завидов. — М. : Лига Разум, 1998. — 448 с.
10. Иларионова, В.А. Судебная автотехническая экспертиза / В.А. Иларионова. — М. : Судэкс, 2001. — 156 с.
11. Пучкин, В.А. Основы экспертного анализа дорожно-транспортных происшествий: База данных. Экспертная техника. Методы решений /
B.А. Пучкин — Ростов н/Д : ИПО ПИ ЮФУ, 2010. -400 с.
12. Pogotovkina, N.S. Motorization in Russia: Challenges and solutions [Электронный ресурс] / N.S. Pogotovkina, Z.V. Almetova, Y.N. Gorchakov, S.A. Kosyakov, V.D. Khegay // International Journal of Appliend Engineering Research / India Publications, 2015 — Режим доступа: http://www.scopus.com/results.
EXPERT ASSESSMENT OF THE TERM
«CROSSROAD»AND THE DEFINITION OF ITS BOUNDARIES
V.A. Gorodokin, Z.B. Almetova, V. D. Shepelev
Abstract. Traffic police statistics show that the most common types of road accidents began hitting vehicles, most of which occur at crossroads of roads. An objective investigation of circumstances traffic incidents and identification the true culprit of the accident is the main task of the persons investigating of
the circumstances of the accident. However, the investigation into circumstances of incident is inextricably linked to a clear understanding where, in what place there was a traffic accident and who of the participants used a preferential right to movement. The solution of these issues is often hampered due to insufficiently precise wording of the terms used in the Rules of the road Russian Federation. In particular, this disadvantage concerns the term «crossroad.» In the various literature devoted to the issues of traffic safety, examples are given that allow to determine the configuration and boundaries of the crossroad. However, the vast majority of the examples based on the simplest form of three — or four-way crossroad. In this work, the authors analyzed requirements and came to the conclusions that the necessity of changing the existing term «crossroad».lwe
Keywords: traffic accident, crossroad, traffic safety, rules of the road, the boundaries of the crossroad.
References
1. GIBDD utverzhdaet, chto vse pokazateli avarij-nosti snizilis’ [The traffic police says that all the rates of accidents declined]. Available at: : http://www.zr.ru/content/news/845054-gibdd-utverzhdaet-chto-vse-pokazateli-avarijnosti-snizilis/.
2. V.A. Gohman i dr. Peresechenie i primykanija avtomobil’nyh dorog [The intersection and junction of roads]. Moscow, Vysshaja. shkola, 1977. 310 p.
3. Pravila dorozhnogo dvizhenija Rossijskoj Fede-racii: po sostojaniju na 21.01.2015 g [Traffic rules of the Russian Federation: at 21.01.2015]. Moscow,
000 IDTR, 2015. 64 p.
4. Konvencija o dorozhnom dvizhenii. Evropejs-koe soglashenie, dopolnjajushhee Konvenciju o dorozhnom dvizhenii, otkrytuju dlja podpisanija v Vene 8 nojabrja 1968 goda (Zheneva 1 maja 1971 goda) [Convention on road traffic. European agreement supplementing the Convention on road traffic opened for signature at Vienna on 8 November 1968 (Geneva,
1 may 1971]. Moscow, Associacija mezhdunarodnyh avtomobil’nyh perevozchikov, 1994. 53 p.
5. Kremenec Ju.A. Tehnicheskie sredstva reguli-rovanija dorozhnogo dvizhenija [Technical means of traffic regulation]. Moscow, Transport, 1981. 252 p.
6. Gorodokin V.A. O nekotoryh problemah bezo-pasnosti peshehodov [About some problems of pedestrian safety]. Modernizacija i nauchnye issledovanija v transportnom komplekse, 2015, no 1. pp. 231 — 237.
7. Suvorov Ju.B. Sudebnaja dorozhno-transportnaja jekspertiza. Sudebno-jekspertnaja ocenka dejstvij voditelej i drugih lic, otvetstvennyh za obespechenie bezopasnosti dorozhnogo dvizhenija, na uchastkah DTP [Court traffic expertise. Forensic assessment of the actions of drivers and other persons responsible for ensuring traffic safety on sections of road accident]. Moscow, Jekzamen, 2003. 208 p.
8. Gorodokin V.A. Pravovye, tehnicheskie i vikti-mologicheskie problemy obespechenija bezopasnosti
na nereguliruemyh peshehodnyh perehodah [Legal, technical and victimological problems of safety on unregulated pedestrian crossings]. Cheljabinsk, Cicero, 2012. 130 p.
9. Selivanov N.A. Rassledovanie dorozhno-transportnyh proisshestvij [The investigation of traffic accidents]. Moscow, Liga Razum, 1998. 448 p.
10. Ilarionova V.A. Sudebnaja avtotehnicheskaja jekspertiza [Judicial autotechnical expertise]. Moscow, Sudjeks, 2001. 156 p.
11. Puchkin V.A. Osnovy jekspertnogo analiza do-rozhno-transportnyh proisshestvij: Baza dannyh. Jekspertnaja tehnika. Metody reshenij [Basis of expert analysis of road accidents: the Database. Expert technique. Solutions]. Rostov na Donu, IPO PI JuFU, 2010. 400 p.
12. Pogotovkina N.S. Motorization in Russia: Challenges and solutions // International Journal of Appliend Engineering Research, India Publications, 2015. Available at: http://www.scopus.com/results.
Городокин Владимир Анатольевич (г. Челябинск, Россия) — кандидат юридических наук, доцент, профессор кафедры «Автомобильный транспорт» ФГАОУ ВО «ЮУрГУ (НИУ)» (454080, г. Челябинск, пр. Ленина, 76, e-mail: gorodok_ vlad@mail. ru).
Альметова Злата Викторовна (г. Челябинск, Россия) — кандидат технических наук, доцент кафедры «Автомобильный транспорт» ФГАОУ ВО «ЮУрГУ (НИУ)» (454080, г. Челябинск, пр. Ленина, 76, e-mail: [email protected]).
Шепелев Владимир Дмитриевич (г. Челябинск, Россия) — кандидат технических наук, доцент кафедры «Автомобильный транспорт» ФГАОУ ВО «ЮУрГУ (НИУ)» (454080, г. Челябинск, пр. Ленина, 76, e-mail: [email protected]).
Vladimir A. Gorodokin (Chelyabinsk, Russian Federation) — Ph. D. in Legal Sciences, Ass. Professor, Department Motor Transport, South Ural state University (national research university) (454080, Lenin av, 76, Chelyabinsk, Russian Federation, e-mail: gorodok_ vlad@mail. ru).
Zlata V. Almetova (Chelyabinsk, Russian Federation) — Ph. D. in Technical Sciences, Department Motor Transport, South Ural state University (national research university)» (454080, Lenin av, 76, Chelyabinsk, Russian Federation, e-mail: zlata. almetova@yandex. ru).
Vladimir D. Shepelev (Chelyabinsk, Russian Federation) — Ph. D. in Technical Sciences of the Department Motor Transport, South Ural state University (national research university) (454080, Lenin av, 76, Chelyabinsk, Russian Federation, e-mail: shepele v1978@mail. ru).
Верховный суд пересмотрит понятие «перекресток» — Российская газета
В Верховный суд подан иск о признании частично недействующим одного пункта Правил дорожного движения. Речь идет о термине «перекресток».
Каждый водитель знает, что зона действия многих знаков распространяется до конца населенного пункта или до ближайшего перекрестка.
Но не всякий водитель в путанице наших дорог и правил четко определит, что считать перекрестком. Как правило, заблуждения на этот счет заканчиваются штрафом. Простейший пример: едет человек по Третьему транспортному кольцу, где знаками установлено ограничение 80 км/ч с «ненаказуемой» скоростью 95 км/ч в крайнем левом, пятом ряду. Примыкает к ТТК некая маленькая улочка. И знаков никаких за ней нет. Да ее вообще сложно заметить из крайнего левого ряда. И продолжает водитель ехать свои 95 км/ч, не подозревая о том, что они стали наказуемыми. Ведь он проехал перекресток. А ограничение «80» действовало именно до этой маленькой улочки. А после нее, как и везде в городе, «60».
Если выезд с прилегающей территории не предназначен для сквозного проезда, действие знака не прерывается
Или ищет водитель место для парковки. Проехал мимо знака «Остановка запрещена», через некоторое время увидел проезд, а за ним никаких знаков нет. Там и остановился. Но в результате был наказан за остановку в зоне действия знака «Остановка запрещена». Оказывается проезд — это вовсе не дорога, а выезд с прилегающей территории. Грубо говоря, из двора. Как отличить одно от другого?
Именно в такой ситуации оказалась юрист Любовь Кобцева. Получив постановление о штрафе, она подала иск в Верховный суд. В нем она просит ВС признать частично недействующим пункт 1.2 Правил дорожного движения в части определения понятия «перекресток».
По ее мнению, нынешняя формулировка не согласуется с тем, что указано в Венской международной конвенции о дорожном движении 1968 года, которую подписала Россия.
Согласно конвенции, термин «перекресток» обозначает любое пересечение на одном уровне, соединение и разветвление дорог. Термин «дорога» означает всю полосу отвода любой дороги или улицы, открытой для движения.
Как сообщил газете «Коммерсантъ» официальный представитель МВД в Верховном суде Гайк Марьян, перекрестком, согласно правилам, является пересечение дорог, а дорога — это «обустроенная или приспособленная и используемая для движения полоса земли либо поверхность искусственного сооружения. Если пересекаются две обустроенные дороги — с разметкой и знаками,- это является полноценным перекрестком, после которого действие знака прерывается. Если же знаков и разметки нет, выезд ведет с прилегающей территории, двора или парковки и не предназначен для сквозного проезда, действие знака не прерывается.
Но вот беда, у нас дворами некоторые населенные пункты можно насквозь проехать. Даже в Москве таких сквозных дворовых проездов масса. И есть выезды из дворов, которые оборудованы разделительной разметкой, бордюрами и знаками лучше, чем иная дорога.
Как пояснили корреспонденту «РГ» в Госавтоинспекции, к терминам Правил дорожного движения претензий нет. Но есть претензии к организаторам дорожного движения, которые должны устанавливать дублирующие знаки в спорных ситуациях. Например, если двор имеет сквозной проезд. Кстати, и ГОСТ о дорожных знаках рекомендует дублировать такие указатели после выездов с прилегающих территорий. Однако это лишь рекомендация, которую не всегда выгодно соблюдать. Лишний штраф казны не портит.
Комментарий
Сергей Смирнов, юрист
Если смотреть с точки зрения прямого толкования, то определения в наших правилах не соответствуют терминам Венской конвенции. Но как на это будет смотреть суд — сказать сложно. У нас международные договоры имеют большую силу, чем законы. Не говоря уже о постановлении правительства, которым утверждаются Правила дорожного движения.
Лев Воропаев, адвокат
Вероятнее всего, Верховный суд не удовлетворит этот иск. Во-первых, ВС не является органом, который уполномочен вносить изменения в нормативно-правовые акты, если тот или иной заявитель считает это целесообразным. ВС лишь проверяет их на соответствие высшим законодательным актам.
Что касается Венской конвенции, то она указывает на то, что договаривающиеся стороны имеют право конкретизировать тот или иной термин. Скорее всего суд сочтет, что правила в части определения термина «перекресток» не противоречат ни российским правовым актам, ни Венской конвенции. Тем более, что ВС уже рассматривал подобный иск и противоречий не нашел.
ГРАНИЦЫ ТОБОЛЬСКОГО ГОРОДСКОГО ОКРУГА
25 января 2010
Приложение 317
к Закону Тюменской области
«Об установлении границ
муниципальных образований Тюменской области
и наделении их статусом муниципального района,
городского округа и сельского поселения»
ОПИСАНИЕ
ГРАНИЦЫ ТОБОЛЬСКОГО ГОРОДСКОГО ОКРУГА
Начальной точкой границы Тобольского городского округа является точка 1, расположенная на стыке границ Овсянниковского, Башковского сельских поселений и Тобольского городского округа в 0,30 км на юг от улучшенной грунтовой дороги.
От точки 1 граница идет на северо-восток, юго-восток и снова на северо-восток протяженностью 12,14 км, огибая садовые участки по северо-западной границе отвода шоссе, до точки 2, расположенной в 1,94 км к северо-востоку от перекрестка автодорог.
От точки 2 граница идет на северо-восток на протяжении 2,90 км, петляя по руслу реки и вокруг железнодорожного полотна, до точки 3, расположенной в 1,94 км на северо-восток от перекрестка дорог.
От точки 3 граница идет в юго-восточном направлении на протяжении 9,99 км сначала по железнодорожному полотну, затем, огибая с запада д. Денисова, с запада и юга д. Соколовка, до точки 4, расположенной на южной границе д. Соколовка на улучшенной грунтовой дороге.
От точки 4 граница идет в северо-восточном направлении на протяжении 3,97 км, огибая с восточной стороны д. Соколовка, пересекая линии ЛЭП, огибая с востока д. Михайловка, далее в северо-западном направлении по северо-западной границе отвода железной дороги на протяжении 0,84 км, затем пересекая ее в северо-восточном направлении, на протяжении 1,85 км до точки 5, расположенной на стыке границ Башковского, Верхнеаремзянского сельских поселений с границей Тобольского городского округа, в 2,33 км к юго-западу от поворота ЛЭП., на протяжении 7,92 км, далее в юго-западном направлении по руслу реки Еловка на протяжении 5,24 км до точки 6, расположенной на стыке границ Верхнеаремзянского, Абалакского сельских поселений и Тобольского городского округа в 1,90 км на юго-восток от поворота шоссе.
От точки 6 граница идет в юго-восточном направлении на протяжении 3,56 км по лесному массиву, затем на юго-запад, пересекая улучшенную грунтовую дорогу, 6,28 км до точки 7, расположенной на стыке границ Абалакского, Прииртышского сельских поселений и Тобольского городского округа в 0,57 км на запад от улучшенной грунтовой дороги.
От точки 7 граница идет в общем юго-западном направлении до шоссе и далее по границе отвода шоссе на протяжении 6,96 км до точки 8, расположенной на юго-западной границе отвода шоссе в 1,15 км на северо-запад от перекрестка автодорог.
От точки 8 граница идет в юго-западном направлении на протяжении 3,39 км, пересекая шоссе, далее вдоль северо-восточного берега реки Иртыш на юго-восток — 3,11 км до точки 9, расположенной в 0,72 км на запад от перекрестка шоссе.
От точки 9 граница идет на юго-запад вдоль шоссе, пересекая его, на протяжении 2,06 км, затем на юго-восток — 3,32 км и далее в северо-восточном направлении протяженностью 3,02 км до точки 10, расположенной на стыке границ Прииртышского, Санниковского сельских поселений и Тобольского городского округа.
От точки 10 граница протяженностью 0,61 км идет на юг и далее в юго-западном направлении на протяжении 5,86 км вдоль озера Саускановского до точки 11, расположенной на стыке границ Санниковского, Ворогушинского сельских поселений и Тобольского городского округа в 0,50 км на юго-запад от пристани.
От точки 11 граница идет в общем северо-восточном направлении, огибая р.п. Сумкино, на протяжении 7,73 км до точки 12, расположенной на реке Иртыш в 2,20 км на север от шоссе.
От точки 12 граница идет на северо-запад на протяжении 11,13 км по середине русла реки Иртыш до точки 13, расположенной на стыке границ Ворогушинского, Овсянниковского сельских поселений и Тобольского городского округа.
От точки 13 граница идет в северо-западном направлении на протяжении 3,41 км по середине реки Иртыш, далее на запад по руслу реки Бееровка и на юго-запад по границе отвода улучшенной грунтовой дороги — 3,37 км, затем на северо-запад вдоль озера Лебяжье протяженностью 3,25 км до точки 14, расположенной на границе отвода железной дороги в 2,80 км на юго-запад от развилки улучшенных грунтовых дорог.
От точки 14 на протяжении 2,64 км граница идет по юго-восточной границе отвода железной дороги в северо-восточном направлении, далее на юго-восток и северо-восток — 2,12 км до точки 15, расположенной в 0,30 км на юг от улучшенной грунтовой дороги.
От точки 15 граница идет в общем северо-восточном направлении, делая петлю по улучшенной грунтовой дороге, на протяжении 4,74 км, далее на северо-запад — 1,27 км и затем вдоль границы отвода улучшенной грунтовой дороги на северо-запад — 4,33 км до точки 16, расположенной в 1,00 км на север от шоссе.
От точки 16 граница идет на северо-восток до реки Иртыш протяженностью 1,86 км, далее по середине реки в северо-западном направлении на протяжении 5,01 км до точки 1.
Настоящее описание границы Тобольского городского округа составлено по топографическим картам 1:100000 издания 1996 года (состояние местности на 1991 год), по опознанным топографическим объектам с учетом материалов землеустройства и лесоустройства и материалов согласования областных границ.
Все расстояния, приведенные в настоящем описании границы, измерены по картам масштаба 1:100000 с точностью до 0,01 км.
Общая протяженность границы Тобольского городского округа составляет 133,27 км.
Экспертная оценка термина «Перекресток» | Городокин
1. ГИБДД утверждает, что все показатели аварийности снизились. — Режим доступа: http://www.zr. ru/content/news/845054-gibdd-utverzhdaet-chto-vse-pokazateli-avarijnosti-snizilis/ (Дата обращения 20.10.2016).
2. Гохман, В.А. Пересечение и примыкания автомобильных дорог : учеб. пособие для вузов / В.А. Гохман и др. — М. : Высшая школа, 1977. -310 с.
3. Правила дорожного движения Российской Федерации: по состоянию на 21.01.2015г. — М. : ООО ИДТР, 2015. — 64 с.
4. Конвенция о дорожном движении. Европейское соглашение, дополняющее Конвенцию о дорожном движении, открытую для подписания в Вене 8 ноября 1968 года (Женева 1 мая 1971 г.). -М. : Ассоциация международных автомобильных перевозчиков, 1994. — 53 с.
5. Кременец, Ю.А. Технические средства регулирования дорожного движения / Ю.А. Кременец, М.П. Печерский. — М. : Транспорт, 1981. — 252 с.
6. Городокин, В.А. О некоторых проблемах безопасности пешеходов / В.А. Городокин, З.В. Альметова // Модернизация и научные исследования в транспортном комплексе. — 2015. — № 1. — С. 231-237.
7. Суворов, Ю.Б. Судебная дорожнотранспортная экспертиза. Судебно-экспертная оценка действий водителей и других лиц, ответственных за обеспечение безопасности дорожного движения, на участках ДТП / Ю.Б. Суворов. — М. : Экзамен, 2003. — 208 с.
8. Городокин, В.А. Правовые, технические и виктимологические проблемы обеспечения безопасности на нерегулируемых пешеходных переходах / В.А. Городокин, А.В Майоров. — Челябинск : Цицеро, 2012. — 130 с.
9. Селиванов, Н.А. Расследование дорожно-транспортных происшествий / Н.А. Селиванов, А.И. Дворкин, Б.Д. Завидов. — М. : Лига Разум, 1998. — 448 с.
10. Иларионова, В.А. Судебная автотехническая экспертиза / В.А. Иларионова. — М. : Судэкс, 2001. — 156 с.
11. Пучкин, В.А. Основы экспертного анализа дорожно-транспортных происшествий: База данных. Экспертная техника. Методы решений / В. А. Пучкин — Ростов н/Д : ИПО ПИ ЮФУ, 2010. -400 с.
12. Pogotovkina, N.S. Motorization in Russia: Challenges and solutions [Электронный ресурс] / N.S. Pogotovkina, Z.V. Almetova, Y.N. Gorchakov, S.A. Kosyakov, V.D. Khegay // International Journal of Appliend Engineering Research / India Publications, 2015 — Режим доступа: http://www.scopus.com/results.
границ | Морфогенез на границе среднего и заднего мозга: на пересечении передачи сигналов Wnt и Fgf
Введение
Граница среднего и заднего мозга (MHB), также называемая истмическим организатором, вызывает интерес биологов, занимающихся развитием, на протяжении десятилетий. Характеризуемый заметным сужением в развивающейся нервной трубке, MHB, расположенный на границе нейромеров среднего и заднего мозга, хорошо известен как центр передачи сигналов, ответственный за формирование паттерна клеточных судеб спереди в среднем мозге и сзади в мозжечке (Wurst и Bally-Cuif, 2001; Raible and Brand, 2004; Dworkin and Jane, 2013).Сужение особенно заметно в спинной нервной трубке и определяет заднюю поверхность среднего мозга и мозжечок заднего мозга. Сужение MHB также разделяет желудочковые области в просвете нервной трубки, при этом желудочек среднего мозга находится впереди сужения, а желудочек заднего мозга — позади него. Таким образом, MHB представляет собой решающую точку разделения в развивающемся головном мозге с характерными морфологическими особенностями, которые имеют решающее значение для нескольких функций MHB: как центр передачи сигналов, как проводник для миграции нейронов и поиска пути аксонов (Volkmann et al., 2010), и как физическое разделение желудочков головного мозга (Lowery et al., 2009). Менее понятна связь между механизмами, ответственными за спецификацию и формирование паттерна MHB, и между сигнальными молекулами, которые обеспечивают активность его сигнального центра, и морфологией сужения, которая неизменно сопровождает развитие MHB у позвоночных. Другими словами, почему всегда имеется сужение нервной трубки в области MHB, и является ли эта морфология причиной или следствием функции MHB?
Механизмы, лежащие в основе спецификации и функций MHB, представляют интерес на нескольких уровнях.Во-первых, исследования на модельных организмах показали, что дефекты в спецификации и формировании паттерна MHB приводят к серьезным нарушениям в мозге, таким как отсутствие среднего мозга, потеря мозжечка и чрезмерный рост тектума среднего мозга (McMahon and Bradley, 1990; Thomas and Capecchi, 1990; Buckles et al., 2004). Во-вторых, достижения в области магнитно-резонансной томографии (МРТ) позволили провести новый анализ пороков развития среднего и заднего мозга человека (Doherty et al., 2013). Эти новые визуализационные исследования выявляют удивительное количество нарушений центральной нервной системы человека, которые, вероятно, вызваны аберрантным формированием паттерна развития, таким как ассоциация септооптической дисплазии с делециями хромосомы 14, которые включают ген формирования нервного паттерна, otx2 (Severino и другие., 2014). Выявление потенциальных причин этих тяжелых заболеваний нервной системы требует глубокого понимания механизмов развития, лежащих в основе развития среднего и заднего мозга.
Как продемонстрировано мутантами мышей и репортерными линиями рыбок данио, MHB специфически расположен внутри домена ранней нервной пластинки, называемого mes / r1 у мыши (Zervas et al., 2004) или доменом заднего мозга среднего мозга (MH) у рыбок данио. (Таллафус и Балли-Куиф, 2003). Эти исследования показывают, что ранний средний мозг (mes) и ромбомер 1 (r1) в передней части заднего мозга генетически согласованы, и MHB определяет точку баланса между этими отделами среднего и заднего мозга.Помимо позиционирования будущего MHB, точка баланса также представляет собой интерфейс между лигандом Wnt, экспрессирующим предшественники заднего среднего мозга, и лигандом Fgf, экспрессирующим предшественники переднего ромбовидного мозга, которые взаимодействуют множеством способов на протяжении спецификации и морфогенеза mes / r1 и MHB. Таким образом, важный вопрос, на который еще нет достаточного ответа, — каково значение интерфейса Wnt-Fgf в MHB для разработки mes / r1?
Морфогенез и формирование паттерна доменов среднего мозга и заднего мозга
Чтобы оценить сложность анализа роли сигнальных семейств Wnt и Fgf в морфогенезе и формировании паттерна MH истмическим организатором, а также для того, чтобы начать идентифицировать процессы, нарушение которых может привести к неврологическому расстройству, полезно сначала получить четкое представление картина того, как MH обретает форму.Модель современного морфологического и молекулярного онтогенеза области MH у рыбок данио показана на рисунке 1.
Рисунок 1 . Морфогенетический и молекулярный онтогенез границы среднего мозга и заднего мозга (MHB) у эмбрионов рыбок данио. Левый столбец: схематические изображения эмбрионов рыбок данио, вид сбоку, на стадиях, указанных слева. Серой штриховкой выделен зачаток мозга. Средний столбец: диаграммы соответствующих морфогенетических движений. Ориентация указана в левой части каждой диаграммы.Примеры каждого морфогенетического свойства можно найти в соответствующих ссылках в правом столбце. fb — передний мозг; мб, средний мозг; hb, задний мозг.
Выбор позиции
Первым критическим шагом в морфогенезе MH является правильное позиционирование зачатка нервной системы на осях тела, так что сигналы пространственно-временного позиционирования могут должным образом «постериоризовать» нервную систему, то есть устанавливать переднезаднюю позиционную информацию после нервной индукции. Позиционирование нервного зачатка частично опосредуется механическим соединением презумптивной нейроэктодермы с инволютивной, перемещающейся кпереди прехордальной пластинкой аксиальной мезодермы посредством сил трения, создаваемых молекулой клеточной адгезии Е-кадгерином, которая на своем цитоплазматическом конце соединена с каноническая эффекторная молекула Wnt, β-катенин (Smutny et al., 2017). В этом исследовании несвязанные эктодермальные клетки перед презумптивной нейроэктодермой «текли» латерально, кзади, а затем медиально. Эти сложные «вихревые» миграции предполагаемых нейроэктодермальных клеток предположительно являются частью конвергентных движений разгибания, которые зависят от неканонической передачи сигналов Wnt / PCP для облегчения медиальной интеркаляции и удлинения передне-задней оси (Heisenberg et al., 2000; Davey and Moens, 2017 ). Что примечательно, так это то, что в своем бурном путешествии презумптивные нейроэктодермальные клетки точно подвергаются сложной молекулярной программе, которая включает апостериоризацию сигналов Wnt и Fgf, исходящих от края бластодермы (обзор в Green et al., 2015; Tuazon and Mullins, 2015), что привело к формированию нервной пластинки с правильным рисунком. Одним особенно примечательным и неизвестным аспектом этого процесса является то, как возникают ранние клеточные ответы на лиганды Wnt и Fgf во время сложных морфогенетических изменений гаструляции.
Несколько линий доказательств предполагают, что лиганды Wnt и Fgf, экспрессируемые в формирующейся параксиальной мезодерме, действуют как морфогены, устанавливая градиенты концентрации или активности, которые генерируют информацию о переднезаднем положении, которая интерпретируется в структурированные судьбы клеток (например,г., Cox and Hemmati-Brivanlou, 1995; МакГрю и др., 1995). Важнейшая функция, в частности лигандов Wnt в поляризации нервной пластинки, была установлена в экспериментах на рыбках данио, Xenopus и цыплятах (Kiecker and Niehrs, 2001; Nordström et al., 2002; Dorsky et al., 2003; Rhinn et al. , 2005). Механизм, с помощью которого устанавливается градуированная активность Wnt, еще не известен, хотя недавние результаты предполагают, что доставка лиганда Wnt через филоподии от предшественников параксиальной мезодермы может быть основным способом транспорта (Stanganello et al., 2015). Это отличается от Fgf в этом контексте, который, как было показано, свободно диффундирует из своего параксиального источника мезодермы и формирует градиент посредством механизма источник-сток (Scholpp and Brand, 2004; Yu et al., 2009).
Важное значение для положения организатора MHB, в частности, имеет активация фактора транскрипции gbx1 в задней нервной пластинке с помощью Wnt8a (Rhinn et al., 2005, 2009), который противодействует независимо активированным otx2 , экспрессируемым в передний и средний мозг (Kurokawa et al., 2012). У рыбок данио эти домены слегка перекрываются на 60% эпиболии, но впоследствии становятся взаимоисключающими на 80% эпиболии (Rhinn et al., 2003), в то время как у мышей изначально существует разрыв между Otx2 и Gbx2, который закрывается (Wurst and Bally-Cuif , 2001). В любом случае существует территория клеток, которая, по-видимому, не связана ни с компартментом среднего, ни с задним мозгом, что согласуется с наблюдаемой вариабельностью в экспериментах по картированию судеб, проводимых до и после гаструляции (Woo and Fraser, 1995).Однако существуют убедительные доказательства того, что к моменту начала сегментации у рыбок данио (Langenberg and Brand, 2005), цыплят (Tossell et al., 2011a, b) и мышей (Sunmonu et al., 2011) MHB представляет собой линию происхождения. рестриктированы и клетки в компартментах среднего и заднего мозга не смешиваются, хотя этот вывод не является бесспорным (напр., Jungbluth et al., 2001). Хотя правильная компартментализация важна для последующего создания пулов нейральных предшественников соответствующего размера в MH, остается открытым вопрос, вызывает ли физический или молекулярный механизм ограничение клонов в MHB.Сообщение от chick предполагает, что имеется сдвиг кзади в положении интерфейса Otx / Gbx, так что он совпадает только с физическим сжатием MHB на более поздних стадиях развития (Hidalgo-Sánchez et al., 2005). У рыбок данио и у мышей, однако, консенсус состоит в том, что начальная граница Otx / Gbx окончательно отмечает будущее сужение MHB до того, как оно станет морфологически видимым. Передача сигналов Notch, которая обычно заставляет клетки принимать такие решения о границах путем усиления небольших стохастических различий в экспрессии генов с последующей сортировкой клеток, участвует в сортировке клеток Otx и Gbx в MHB у цыплят (Tossell et al., 2011б). Сообщения от мышей и medaka предполагают, что др. Генетические взаимодействия могут уточнять границу, поскольку Gbx2 взаимодействует с репрессорами Groucho и может напрямую конкурировать с активаторами транскрипции POU Otx2 (Heimbucher et al., 2007; Inoue et al., 2012). Межклеточные сети актиномиозина, которые, как было показано, управляют морфогенезом, например, во время инвагинации мезодермы у Drosophila , также могут играть физическую роль в ограничении клонов в MH (Kasza and Zallen, 2011) или, возможно, региональные изменения в кортикальных клетках, вызывающих натяжение актина. для сортировки по ту или иную сторону границы (Heisenberg and Bellaïche, 2013).
Активация
Вскоре после фазы позиционирования во время гаструляции набор генов MHB активируется в отдельных доменах вокруг границы otx2 / gbx , поскольку нервная пластинка подвергается нейруляции с образованием нервной трубки. Экспрессия wnt1 кпереди и fgf8a кзади от предполагаемого MHB (что все еще не является морфологически очевидным) усиливают интерфейс otx2 / gbx , в то время как her5 , eng2a и pax2a выражаются с обеих сторон . граница (Rhinn, Brand, 2001; Buckles et al., 2004). Какие молекулы и / или силы активируют этих основных членов более обширной генетической программы MHB, остается плохо изученным, и, что удивительно, эта программа активации может происходить в отсутствие по крайней мере частей механизма позиционирования (Su et al., 2014). После активации специфическая роль каждого гена в стимулировании последующего развития внутри MH (помимо предоставления пространственных сигналов) также не совсем понятна, хотя некоторые компоненты программы MHB, по-видимому, играют роль как в спецификации судьбы, так и в морфогенезе внутри MH (Dworkin и Джейн, 2013).Напр., her5 , как известно, ингибирует нейрогенез во время сегментации (Tallafuss and Bally-Cuif, 2003; Ninkovic et al., 2005) и впоследствии способствует идентичности нервных стволовых клеток у взрослых рыбок данио (Chapouton et al., 2006).
Однако создание интерфейса сигнализации Wnt / Fgf, безусловно, имеет решающее значение для развития MH. Мыши Wnt1 — / — и Fgf8 — / — не могут развить всю область MH (McMahon and Bradley, 1990; Chi et al., 2003). У рыбок данио потеря нескольких повторяющихся Wnts ( wnt3 , wnt3a , wnt1 и wnt10b ) воспроизводит аналогичный фенотип (Lekven et al., 2003; Buckles et al., 2004) и рыбок данио fgf мутант ace лишен сужения мозжечка и MHB, хотя средний мозг присутствует, но неполяризован, что приводит к аберрантным ретинотектальным проекциям (Picker et al., 1999). Fgf8 был признан наиболее важной «организующей молекулой» на основании результатов имплантации пропитанных Fgf8 шариков в сайты кпереди и сзади от MHB.В этих экспериментах Fgf8 был достаточен для индукции тектальных и мозжечковых структур и лежащих в основе границ Otx / Gbx, тогда как подобные эксперименты для Wnt1 не показали значительного изменения паттерна окружающих тканей (Martinez et al., 1999). В самом деле, никакой прирост функционального анализа для всех других основных молекул MHB в каком-либо организме не дал таких поразительных результатов. Однако исследование, в котором Otx2 и Fgf8 были одновременно сбиты с ног, поставило под сомнение идею, что Fgf8 необходим для формирования паттерна судеб клеток в MHB.Foucher et al. (2006) показали, что в отсутствие Fgf8, если уровни Otx2 были истощены, нейроны мозжечка были способны успешно дифференцироваться, хотя морфология MHB была аномальной у этих эмбрионов. Недавний анализ otx; gbx; fgf эмбрионов также указывает на то, что сильная дифференцировка мозжечка нуждается в Fgf (Su et al., 2014).
Во время активации программы MHB продолжается процесс первичной нейруляции, при котором нервная пластинка сливается с дорсальной срединной линией (Lowery and Sive, 2004).Во время этого процесса медиально-латеральная организация нервной пластинки трансформируется в вентрально-дорсальную ориентацию (Schmitz et al., 1993). Стоит отметить, что, хотя впоследствии развивающееся сужение MHB изучается в первую очередь по отношению к оси A / P, оно неоднородно по оси D / V нервной трубки, что может отражать ступенчатую или неоднородную активность Wnt / Fgf вдоль ось D / V и интеграция с активностью передачи сигналов дорсовентрального паттерна (Lekven et al., 2003; Puelles et al., 2003). Т.о., литература по MHB в основном сосредоточена на активности организаторов в крыловидной области MH, при этом относительно мало известно о базальном покрытии.
Вскоре после формирования нервной трубки и начала миграции нервного гребня сокращение MHB становится видимым морфологическим признаком, поскольку начинают формироваться желудочки мозга. У рыбок данио сокращение MHB требует укорочения клеток и последующего ламинин-зависимого базального сжатия небольшого кольца клеток на границе (Gutzman et al., 2008). Изменения формы клеток, участвующие в морфогенезе констрикции MHB, требуют немышечного миозина II, и недавние результаты показывают, что укорочение клеток, необходимое для сжатия MHB, является следствием временной регуляции кальциевой киназы легкой цепи миозина (Gutzman and Sive, 2010; Gutzman et al. ., 2015; Саху и др., 2017).
Техническое обслуживание
После того, как нервная трубка сформирована и началось сужение MHB, генетическая программа внутри MH впоследствии переходит в поддерживающую фазу, сопровождаемую постоянным изменением формы ткани головного мозга и желудочковой системы, а также производством спинномозговой жидкости, которая сама может вносить свой вклад. к регуляции MHB (Parada et al., 2005; Гато и Десмонд, 2009). Вычислительное моделирование и эксперименты на цыплятах указывают на важность дифференцированной миозин-опосредованной сократимости для создания геометрии желудочков мозга и предполагают, что стратегии могут отличаться от компартмента к компартменту в зависимости от конечной судьбы соединения, поскольку некоторые из них являются лишь временными структурами (например, границы ромбомеров) в то время как другие, такие как сужение MHB, сохраняются как структуры в мозге взрослого человека, которые должны сопротивляться возрастающему давлению жидкости из желудочковой системы (Filas et al., 2012). Одну функцию, которую может выполнять сужение MHB, таким образом, является точкой перехода между различными программами морфогенеза переднего и заднего желудочков мозга, сходящимися на границе. Такая структура должна поддерживать клеточную адгезию на границе до тех пор, пока морфология желудочков мозга не будет установлена с обеих сторон, чтобы предотвратить неправильную спецификацию окружающих тканей. Сужение может также опосредовать синхронизацию передачи сигналов между передним и задним отделами мозга в случае сигнальных молекул, секретируемых в спинномозговую жидкость.Подобные явления не беспрецедентны; например, у эмбрионов мыши хорошо известно, что лево-правая асимметрия нарушается направленным на реснички потоком жидкости в узле, хотя неизвестно, является ли сигнал, опосредованный однонаправленным потоком жидкости, механическим или химическим по своей природе (Yoshiba and Хамада, 2014). Желудочки мозга имеют реснички, а реснички рыбок данио в развивающемся конечном мозге, как было установлено, направляют миграцию нейронов (Kishimoto et al., 2011). Некоторые из таких механизмов могут объяснять эволюцию стратегии закрытой первичной нейруляции, наблюдаемой у рыбок данио по сравнению с складкой нервной трубки, наблюдаемой у других позвоночных.
В фазе поддержания появляются несколько субрегионов MH, которые выполняют свои собственные морфогенетические программы в ожидании нейрогенеза. В среднем мозге покров зрительного нерва формируется за счет образования плотного слоя клеток, называемого периферическим слоем среднего мозга (PML), в котором находятся медленные клетки-предшественники нейронов, которые дадут начало столбцам нейронов, организованных путем чередования экспрессии протокадгерина, которые заселяют больше anterior tectum корковым способом (Recher et al., 2013; Купер и др., 2015; Rapaciolii et al., 2016). Кзади от MHB формируются ромбическая губа мозжечка и зоны желудочков, из которых позже образуются гранулы и клетки-предшественники Пуркинье, соответственно, до того, как их нервные производные организуются в дорсовентральные слои и медиолатеральные компартменты, которые обеспечивают основу цепи мозжечка (Hashimoto и Hibi, 2012; Millen et al., 2014). MH tegmentum почти не имеет явных морфологических ориентиров, за исключением относительно неглубокого сужения, но его правильное формирование паттерна и морфогенез имеют решающее значение для правильного формирования серотонинергических и холинергических ядер, участвующих в важных поведенческих функциях (Parker et al., 2013).
Правильное создание и поддержание каждой из вышеупомянутых про-нейральных субрегионов MH требует как минимум петли обратной связи передачи сигналов Wnt и Fgf, чтобы установить надлежащие уникальные молекулярные и механические микросреды (Carletti and Rossi, 2008). У мутантов рыбок данио ace (fgf8a) активируется экспрессия wnt1 и нескольких других генов в пограничной области, включая her5 , pax2a и eng2 / 3 , но их экспрессия исчезает как Fgf-зависимая обратная связь. терпит неудачу в раннем и среднем сомитогенезе (Reifers et al., 1998). Сходным образом, объединенная потеря wnt1 / wnt10b / wnt3a у рыбок данио приводит к временной экспрессии fgf8a , pax2a и eng2 / 3 в раннем MHB (Buckles et al., 2004). Мы недавно обнаружили, используя живую многофотонную визуализацию (Гиббс и др., 2014a), что мутанты ace образуют сужение, которое не может должным образом созревать в фазе поддержания (рис. 2), что является морфологическим переходным результатом молекулярной быстротечности (Гиббс и др. ., 2013).Неспособность сжатия продолжать морфогенез в поддерживающей фазе происходит из-за аберрантного клеточного поведения в двух группах клеток. Путем визуализации трансгенной репортерной линии wnt1 (Gibbs et al., 2014b) на фоне ace (fgf8a) мы идентифицировали одну группу клеток, которая не может поддерживать экспрессию wnt1 в задней части среднего мозга и впоследствии координировать собственно морфогенез PML и граничной покрышки, и др. группа, которая неспособна подавить экспрессию wnt1 в дорсальной части r1, чтобы правильно специфицировать пластинку мозжечка (Figures 2, 3).Это наблюдение, основанное на идентификации отдельных клеток, подтверждает предыдущие сообщения о трансформации перешейка / мозжечка в тектальную молекулярную идентичность презумптивного мозжечка, которая происходит при генетическом репрограммировании во время поддерживающей фазы (Jászai et al., 2003; Gibbs, 2014 ), хотя при визуализации в реальном времени мы наблюдали, что это перепрограммирование, вызванное отсутствием fgf8a , не препятствует предыдущему инициированию морфогенеза MHB во время фазы активации. Т.о., механизм, независимый от положений fgf8a и , и инициализирует эту физическую границу, тогда как fgf8a- зависимый механизм (прямо или косвенно) поддерживает свой продолжающийся морфогенез.
Рисунок 2 . Зависимый от региона ответ репортера wnt1 на фоне ace (fgf8a) . (A) 3-D реконструкция, созданная с использованием проекции максимальной интенсивности от автофлуоресценции ткани (с удаленными участками верхней пластины), выявляет формирование истмического сужения (желтые стрелки), которое не может созреть без fgf8a . (B) wnt1 линия присутствует, но неправильно поляризована в эмбрионах ace .Во время нормального развития линия wnt1 увеличивает экспрессию wnt1 , что может быть визуализировано по усилению репортерного сигнала eGFP на границе MHB. (C) Линия wnt1 в дорсальном нейроэпителии обычно выключает экспрессию wnt1 в переднем заднем мозге, что отражается в снижении репортерной интенсивности (измеряется по профилям, отмеченным зелеными линиями с течением времени). Красные стрелки указывают на клетку среднего мозга непосредственно перед сужением MHB, а желтые стрелки указывают на соседнюю клетку позади сужения.Синяя стрелка показывает предполагаемый периферический слой среднего мозга (PML). У эмбрионов ace (fgf8a) сжатие расслабляется и соседние клетки в r1 начинают повторно экспрессировать wnt1 , как показано увеличением интенсивности репортера, отражая трансформацию мозжечка в тектальную. Базальное сужение граничных клеток с наивысшей репортерной интенсивностью wnt1 происходит в обоих случаях, однако презумптивный PML не может сформироваться у ace (fgf8a) эмбрионов. (D) Линия wnt1 в вентральном нейроэпителии обычно претерпевает укорочение клеток и сжимается до узкого кольца клеток перед физическим сжатием MHB (черные маркеры), в то время как у ace (fgf8a) эмбрионов эта реорганизация не может произойти. происходит и wnt1 больше не выражается.Масштабная линейка = 100 мкм.
Передача сигналов Wnt и Fgf также может играть более прямую роль в формировании MH во время нейрогенеза. Как упоминалось ранее, на ранних стадиях формирования MHB нейрогенез активно ингибируется с помощью her5 у рыбок данио (Tallafuss and Bally-Cuif, 2003), но впоследствии нейроны рождаются, когда экспрессия her5 отступает к узкому кольцу в месте сужения. Недавно было предложено, что Wnt1 опосредует синхронизацию нейрогенеза в среднем мозге, управляя экспрессией Fgf8 на границе и постепенно подавляя ее от границы, индуцируя экспрессию Sprouty, так что Fgf-зависимый her5 также отступает (Dyer et al., 2014). Wnt1 также может действовать, способствуя идентичности нервных стволовых клеток в дорсальной части среднего мозга и MHB (Miyake et al., 2012; Lin and Lee, 2016), возможно, регулируется Fgf3 / 8-зависимой передачей сигналов Fgf22 (Miyake and Itoh, 2013) и может вносят вклад в формирование MH, регулируя цитоскелет во время наведения аксонов (Ciani and Salinas, 2005). В заднем мозге дифференциация уникальных идентичностей ядер покрышки происходит в пространственно-временных волнах, исходящих от верхней ромбической губы. Недавно было показано, что эти миграции сохраняются у мышей и рыбок данио, при этом дискретные популяции Wnt1 в верхней ромбической губе последовательно мигрируют вперед к MHB и поворачиваются вентрально к своим конечным положениям в тегментуме заднего мозга (Volkmann et al., 2010). Передача сигналов Fgf9 / Fgfr2 важна для дифференцировки глиальных клеток Бергмана в мозжечке мышей (Meier et al., 2014), клеточного типа, консервативного в мозжечке рыбок данио (хотя рыбки данио, по-видимому, не имеют fgf9 , эта функция может можно отнести к другому избыточно функционирующему Fgf; Bae et al., 2009).
Морфогенетические роли в передаче сигналов Wnt и Fgf во время формирования констрикции
Сигнальные пути Wnt и Fgf представляют собой обширные основные пути развития, которые играют разнообразные контекстно-зависимые роли.В этом разделе мы дополнительно исследуем концепцию, что сигнальная петля Wnt / Fgf необходима для собственно морфогенеза MH, и обсуждаем потенциальные точки перекрестного взаимодействия между этими сигнальными путями и клеточной адгезией и цитоскелетным аппаратом, основываясь на исследованиях как в MH, так и в других системах.
Влияние модуляции передачи сигналов Wnt / Fgf на морфогенез MH
Потеря fgf8a в MH у мутантов ace и последующая модуляция экспрессии wnt1 внутри линии wnt1 (рисунки 2, 3) приводит к неспособности клеток вблизи сужения MHB выполнять определенные аспекты нормальный морфогенез, включая укорочение клеток, сохранение адгезии по средней линии, правильное формирование желудочков мозга и организацию презумптивного ПМЛ (Гиббс, 2014).Тем не менее, существует некоторый механизм (возможно, Wnt-зависимый) из фазы позиционирования или активации, который инициирует временное сжатие.
Рисунок 3 . Нарушение адгезии у ace (fgf8a) эмбрионов и регионально зависимый ответ клонов wnt1 . (Слева) Первоначально широкая экспрессия wnt1 обычно уточняется в дорсальном среднем мозге и переднем среднем мозге в домене заднего мозга (MH), помогая поддерживать fgf8a -зависимую адгезию в дорсальной MH.(Справа) wnt1 Экспрессия в задней части среднего мозга (за исключением дорсальной полосы) теряется в ace (fgf8a) , тогда как экспрессия wnt1 в дорсальной полосе r1 реактивируется во время трансформации мозжечка в тектальную. Срединная адгезия в ЗГ не поддерживается во время морфогенеза желудочков головного мозга.
Из изображений в реальном времени мы можем сделать вывод, что адгезия по средней линии и релаксация сужения отделены друг от друга в фенотипе ace ( fgf8a) , поскольку момент времени, в который началось отслеживание на рис. r1, но ориентация клеток на границе медленно менялась со временем от перпендикулярной к наклонной (противоположной нормальной ориентации) относительно оси A / P (Gibbs, 2014).Мы интерпретируем эти изменения в ориентации как отражение другого механизма, который вносит вклад в фенотип ace (fgf8a) независимо от потери адгезии. Одна из возможностей — опосредованная миозином стадия эпителиальной релаксации, аналогичная механизму, используемому для создания временных сужений между ромбомерами заднего мозга (Gutzman and Sive, 2010), поскольку было показано, что немышечные миозины важны для образования сужений MHB (Gutzman et al. , 2015). Без надлежащего натяжения эпителия задней части среднего мозга базально-сужающие клетки могут быть не в состоянии механически стабилизироваться окружающими тканями, что приводит к потере правильной морфологии границ.Мы наблюдали, что wnt1 экспрессирующие клетки в заднем дорсальном среднем мозге организуются в простой эпителий в MHB, который совпадает с предполагаемым PML. Возможно, эта организация обеспечивает локальную механическую жесткость, которая позволяет базальному сужению пограничных клеток приводить к движению в переднем и заднем направлении. Или, возможно, базальная мембрана этого эпителия взаимодействует с базальной мембраной r1, чтобы стабилизировать область во время базального сужения.
Мы также отслеживали аналогичную динамику в вентральной области MH, показанной на рисунке 2D, хотя ослабление сигнала на этих глубинах визуализации затрудняло отслеживание отдельных клеток (Gibbs, 2014). Мы заметили, что линия wnt1 , первоначально распространенная через границу, впоследствии была сжата в более узкую область по мере увеличения угла границы (черные маркеры). Было неясно, было ли такое поведение результатом базального сужения. Напротив, у эмбрионов ace (fgf8a) линия wnt1 , первоначально охватывающая границу, не смогла укорачиваться, сжиматься и организовываться в кольцо клеток, хотя их ориентация, казалось, перешла от перпендикулярной к наклонной, возможно, как результат сил, возникающих в результате нарушения сужения дорсального нейроэпителия.Исходя из этого анализа, казалось бы, существуют разные механические силы, действующие на сужение перешейка вдоль дорсовентральной оси трубки во время сомитогенеза, что потенциально может привести к различным стратегиям поддержания механической целостности границы для правильного формирования окружающей ткани.
Ранее повышенные уровни гибели клеток наблюдались в дорсальном r1 у ace мутантов во время среднего сомитогенеза (Reifers et al., 1998) и приписывались апоптозу после изменения идентичности r1-мезэнцефалии.Сходный фенотип Fgf8 — / — наблюдался у мышей (Chi et al., 2003). Хотя такое явление может потенциально объяснить потерю адгезии, которую мы наблюдаем в дорсальном r1, мы не наблюдали какого-либо увеличения сигналов автофлуоресценции, которые могут указывать на такие события, происходящие у ace (fgf8a) эмбрионов. Вместо этого мы утверждаем, что fgf8a играет роль в поддержании адгезии во время нормального развития. Мы также наблюдали у эмбрионов ace (fgf8a) , что открытие заднего мозга инициируется нормально, но распространяется вперед в MH, возможно, из-за неспособности поддерживать генетическую программу MHB без fgf8a (Gibbs, 2014).Недостаточная передача сигналов Fgf может, таким образом, делать дорсальный MH компетентным к сигналу, исходящему от границы r1 / r2, который запускает открытие желудочков заднего мозга (Gutzman and Sive, 2010). Интересно, что пределы, до которых сужается изначально широкий домен fgf8a (первоначально охватывающий r1-r4 и суженный до дискретных доменов в r1, вентральном r2 и r4; Reifers et al., 1998), отражают любопытную последовательность открытия желудочков заднего мозга. в более задних ромбомерах. У рыбок данио желудочки открываются сначала на дорсальной границе r1 / r2 (Gibbs, 2014), за ними следуют небольшие отверстия на границах r3 / r4 и r4 / 5 (Gutzman and Sive, 2010).Время разделения r4, описанное Gutzman and Sive (2010), происходит, по-видимому, в то же время, когда экспрессия fgf8a теряется в r4 (Reim and Brand, 2002). Поскольку домены переднего мозга экспрессии fgf8a также соответствуют областям, которые формируют морфологию желудочков переднего мозга, а большой домен fgf8a на границе otx / gbx эмбрионов с потерей функции соответствовал аномально длительному сжатию, при котором клетки, казалось, не могли должным образом функционировать. расслаиваются на их апикальном интерфейсе (Su et al., 2014), разумно предположить, что fgf8a может быть необходимым фактором для поддержания адгезии на границах желудочков мозга.
Рисунок 4 суммирует эффекты модуляции передачи сигналов Wnt / Fgf на поведение морфогенных клеток в MH во время поддерживающей фазы формирования. Полный нокдаун fgf8a и временные низкие уровни экспрессии wnt1 в эмбрионах ace приводят к фенотипу адгезии, противоположному тому, который наблюдается, когда экспрессия fgf8a инициируется неограниченным образом во всем MH и впоследствии активируется в конце . Передача сигналов wnt1 в эмбрионах с потерей функции otx / gbx (Su et al., 2014). Роль, которую играет fgf8a в поддержании адгезии, вероятно, косвенная. Потеря адгезии по средней линии резко прогрессирует в соответствии с потерей паттернов экспрессии eng2 и pax2a в ace в MHB (Reifers et al., 1998) и eng2 , например fgf8a , отсутствует. в r4, когда там открывается IV желудочек. Было бы интересно изучить эти паттерны подробно, чтобы увидеть, экспрессируются ли eng2 или pax2a больше в клетках на средней линии и потенциально опосредуют ли поддержание адгезии. wnt1 , eng1.b , pax2a и il17rd перекрываются с fgf8a в этом регионе и могут быть генами-кандидатами, которые помогают опосредовать это fgf8a зависимое разделение в MH. Время экспрессии wnt1 , независимо от того, происходит ли оно только во время фазы активации (как в ace (fgf8a) ) или во время фазы поддержания (как в otx / gbx потеря функции), может быть более важным для поведения клеток. такие как инициация сужения, укорочение клеток и образование PML в дополнение к ограничению fgf8a спереди.
Рисунок 4 . Модуляция передачи сигналов Wnt / Fgf и влияние на морфологию MH. (Слева) Сводка доменов экспрессии wnt / fgf , морфологии MH и морфогенетического поведения клеток у мутантов ace . (В центре, справа) Та же модель показана для эмбрионов дикого типа и otx / gbx эмбрионов с потерей функции. Рисунок составлен на основе данных, найденных в Su et al. (2014). Дефицитное поведение клеток обозначается знаком минус (-), а поведение сверхактивных клеток обозначается знаком плюс (+).
Перекрестные помехи Wnt / Fgf с клеточной адгезией и цитоскелетными механизмами
На всех этапах позиционирования, активации и поддержания образования MH установлено, что каноническая и неканоническая сигнальная активность Wnt, сигнальная активность Fgf, а также точная модуляция клеточной адгезии, полярности и подвижности необходимы для правильного формирования зародышевые листки, нейральный зачаток и, следовательно, MH. Эти молекулярные взаимодействия, которые могут определять поведение клеток, ведущее к этим морфогенетическим изменениям, суммированы на Рисунке 5.Внутри MH, вероятно, будут пространственно-временные изменения компетенций к этим взаимодействиям, но поскольку они не были точно определены в этой конкретной биологической системе, они показаны вместе в одной ячейке как возможные пути для дальнейшего исследования.
Рисунок 5 . Возможные пути передачи сигналов Wnt / Fgf, способствующие образованию MH.
Каноническая передача сигналов Wnt уже давно связана с накоплением пула цитоплазматического бета-катенина путем ингибирования его деградации, так что бета-катенин может перемещаться в ядро и регулировать гены-мишени с сайтами связывания TCF / LEF (Moon et al., 2004). Однако передача сигналов как Wnt, так и Fgf может ингибировать GSK3, первичный компонент комплекса разрушения, который модулирует уровни бета-катенина (Dailey et al., 2005). Таким образом, тщательный баланс передачи сигналов Wnt / Fgf может влиять на гены-мишени Wnt во время развития MH, такие как snail (Yook et al., 2005) и twist (Klymkowsky et al., 2010), zic2a / 5 ( Nyholm et al., 2009), cyclinD (Megason and McMahon, 2002), stmn4 (Lin and Lee, 2016) и dpagt1 (Varelas et al., 2014), гены, которые, как известно, влияют на пролиферацию клеток, формирование дорсолатеральной шарнирной точки (DLHP) и адгезию. Классическая передача сигналов Fgf также может быть важной в спецификации судьбы глии Бергмана в мозжечке (Meier et al., 2014). Передача сигналов Fgf / MAPK может также активировать белки Sox, которые могут ингибировать взаимодействия beta-catenin / TCF / LEF, а также активировать Snail, который способствует эпителиально-мезенхимальному переходу (EMT) путем ингибирования кадгеринов (Dailey et al., 2005).
Перекрестные помехи между передачей сигналов Wnt и опосредованной бета-катенином клеточной адгезией посредством связывания с кадгеринами были областью текущих исследований, выявивших большое количество контекстно-зависимых точек взаимодействия, в которых передача сигналов Wnt может модулировать клеточную адгезию и наоборот посредством перемещения клеток. бета-катенин между цитоплазматическим пулом, который может стать ядерным эффектором, и мембранным пулом, который взаимодействует с цитоскелетом (Heuberger and Birchmeier, 2010).Как эти взаимодействия могут способствовать формированию сужения MHB и других регионов MH, неизвестно. N-cadherin рыбок данио ( cdh3 ) экспрессируется во всей нервной системе, а мутант cdh3 parachute имеет значительную потерю адгезии по средней линии в нервной трубке (Lele et al., 2002). Подавление cdh3 осуществляется при миграции клеток нервного гребня Wnt-зависимым образом (Piloto and Schilling, 2010). E-кадгерин рыбок данио ( cdh2 ) не экспрессируется в нервной трубке до некоторого времени после стадии 16 сомитов и экспрессируется в предполагаемом MHB через 24 hpf в области, которая, по-видимому, перекрывается с экспрессией Wnt около средней линии (Babb et al. ., 2001). E-кадгерин может быть уникально ответственным за поддержание клеточной адгезии, чтобы помочь стабилизировать цитоскелетные перестройки по средней линии в области MH или во время образования PML. Один пример, связывающий передачу сигналов Fgf с морфогенетическим ремоделированием цитоскелета, был предложен для работы посредством передачи сигналов Fgfr-Ras-MAPK в формировании сенсорной системы боковой линии у рыбок данио (Harding and Nechiporuk, 2012). В этом исследовании авт. Обнаружили, что передача сигналов Ras-MAPK, активируемая с помощью Fgfr, необходима для образования розеток за счет локализации Rho-ассоциированной киназы (Rock) на апикальной поверхности, чтобы управлять ее сжатием.Передача сигналов Fgf, как было установлено, также играет роль в формировании отических пузырьков, которое требует апикального сужения, опосредованного локальным увеличением актина. В отических пузырьках передача сигналов Fgf активирует фосфолипазу-C (PLC), которая запускает неканоническую активность миозина-II (Sai and Ladher, 2008). Считается, что классически миозин-II действует вдоль актиновых филаментов, чтобы способствовать сокращению, однако при фосфорилировании с помощью PLC миозин-II способствует деградации базального актина (что приводит к обогащению апикального актина и инвагинации отических чашечек).Передача сигналов Wnt также участвует в ремоделировании цитоскелета посредством пути планарной клеточной полярности и, возможно, также канонических путей передачи сигналов (Lapebie et al., 2011). Как только нейрогенез начинается в области MH, возможно, что передача сигналов Wnt / Fgf может модулировать миграцию нейронов (Knosp et al., 2015). Wnt и Fgf участвуют в изменениях адгезии эпителиальных клеток в нейрогенных краниальных плакодах (Lassiter et al., 2014), и было показано, что новые рецепторы Wnt Ryk и Ror могут взаимодействовать с цитоскелетом, способствуя ведению аксонов (Clark et al., 2012).
Баланс внутри- и межклеточного кальция является еще одним интересным кандидатом-мишенью, связывающим передачу сигналов Wnt / Fgf с клеточной адгезией и динамикой цитоскелета (Kim et al., 2011; Tsai et al., 2015). Передача сигналов Fgf может способствовать высвобождению внутриклеточного кальция и влиять на организацию цитоскелета через кальций и кальмодулин-зависимые протеинкиназы (Schlessinger, 2000), функция, которая также может выполняться Wnt (Babb et al., 2001; Cohen et al., 2008) и может быть комбинаторным в формировании перетяжки MHB.Выводы Gutzman et al. кальциевых транзиентов, которые, по-видимому, управляют миозин-зависимым укорочением клеток в задней части среднего мозга, подчеркивают потенциал такой морфогенетической роли Wnt / Fgf в MH (Gutzman et al., 2015; Sahu et al., 2017). Кроме того, то, как внешние и внутренние физические силы, запускающие пути механотрансдукции, могут пересекаться с путями формирования паттерна ткани, продолжает оставаться активной областью исследований (Heisenberg and Bellaïche, 2013).
Как показано на рисунке 4, поведение нескольких клеток в области MH во время фазы поддержания, по-видимому, зависит от передачи сигналов Wnt / Fgf, но исследования только начинают определять, какие молекулярные компоненты клеточной адгезии, цитоскелета или механотрансдукции ответственны за конкретные поведения и еще не перечислили прямые пути ниже Wnt или Fgf, которые действуют в этой области.Потеря передачи сигналов Fgf8a и связанная с этим неспособность поддерживать экспрессию wnt1 у мутантов ace (fgf8a) приводит к неспособности клеток должным образом укорачиваться при сужении, аберрантному раздуванию желудочков и морфологии в задней части среднего мозга, потере адгезии по средней линии в сжатие и неспособность PML начать формироваться; тем не менее, базальное сужение граничных ячеек действительно происходит. Расширенная передача сигналов Wnt / Fgf (при этом передача сигналов Wnt не становится активной до фазы поддержания) у мутантов gbx1 / 2 с нокдауном otx2 приводит к большому сужению из-за чрезмерного прилегания к средней линии, очевидно нормальному раздуванию желудочков и неизвестному влиянию на укорочение клеток. , базальное сужение и образование PML.На рисунке 5 показаны потенциальные пути, связывающие передачу сигналов Wnt / Fgf и возможные нижестоящие эффекторы этого клеточного поведения, которые продолжают требовать дальнейшего изучения, поскольку остается трудным изучить или визуализировать, как множественные входные сигналы передаются вторичными молекулами и нижестоящими эффекторами для регуляции транскрипции генов, цитоскелета. динамика, адгезия клеток и последующее поведение клеток in vivo .
Новые инструменты для работы со старым модельным органайзером
Несмотря на все, что мы знаем о передаче сигналов Wnt и Fgf и развитии MHB, важные вопросы, такие как формирование паттерна ткани посредством передачи сигналов клеток, пересекаются с генерацией морфогенетической силы и изменением формы клеток, остаются без ответа.Одним из препятствий на пути к значительному дальнейшему прогрессу было отсутствие соответствующих технологий, позволяющих визуализировать как идентичность клеток, так и изменения цитоскелета. Классически морфогенез и формирование паттернов изучались относительно разрозненно из-за технологических ограничений, касающихся масштаба и точности генетических и эмбриологических манипуляций и молекулярных меток, пространственно-временного и спектрального разрешения систем визуализации, сложности автоматизации сложных задач обработки изображений и минимального сотрудничества. среди биологов развития, физиков и инженеров.Все эти ограничения явно подчеркиваются пробелами в понимании механизмов формирования паттерна и морфогенеза области MH. В этом разделе мы кратко обсуждаем последние достижения в области визуализации и технологий обработки изображений, которые, как мы надеемся, помогут собрать более пространственно-временную модель формирования паттерна и морфогенеза MH.
Изображения
Изображения, представленные на Рисунке 2, были получены с помощью самодельной системы микроскопии ультракоротких импульсов (UPM), сконфигурированной для визуализации изображений интенсивности с использованием двухфотонно возбужденной флуоресценции.В нашей системе UPM импульсы длительностью менее 10 фемтосекунд от титан-сапфирового генератора с пассивной синхронизацией мод передаются двухосным гальванометрическим сканером в вертикальный микроскоп. Эмбрионы рыбок данио помещали в лунки с агарозой и погружали для связывания с иммерсионными объективами. Вертикальная геометрия удобна для совмещения интересующей области эмбриона вручную с оптической осью микроскопа. Генерируемая двухфотонная возбужденная флуоресценция собирается в геометрии обратного отражения объективом микроскопа и направляется в фотоумножительные трубки для подсчета фотонов для визуализации изображения.В этой конфигурации наша система UPM представляет собой точечное сканирование, при котором изображения визуализируются в цифровом виде попиксельно (Гиббс и др., 2014b).
Экспериментальная конфигурация нашей системы UPM с точечным сканированием концептуально аналогична системам лазерной сканирующей конфокальной микроскопии (LSCM), и сравнения между LSCM и двухфотонной лазерной сканирующей флуоресцентной микроскопией (2PM) хорошо обсуждались (Gao et al., 2012 ). Здесь мы подчеркиваем два момента: генерацию сигнала и фотообесцвечивание. Сигнал флуоресценции, используемый в LSCM, генерируется после линейного поглощения падающих фотонов; линейное поглощение может быть подтверждено линейной зависимостью между интенсивностью падающего лазера и сигналом флуоресценции.Действительно, «однофотонная» флуоресценция — это легко наблюдаемое явление, так как сигнал генерируется по всему пути луча возбуждения внутри образца. Таким образом, в LSCM перед детектором размещается точечное отверстие, конфокальное с плоскостью объекта, чтобы различать расфокусированный сигнал, чтобы можно было визуализировать тонкие оптические изображения.
Сигнал флуоресценции, используемый в 2PM, генерируется после нелинейного поглощения падающих фотонов, то есть одновременного поглощения двух фотонов. Двухфотонное поглощение может быть проверено квадратичным соотношением между интенсивностью падающего лазера и сигналом флуоресценции.Эта нелинейная зависимость между интенсивностью падающего лазера и флуоресценцией проявляется в ограничении генерации сигнала фокусом луча. Таким образом, в 2 часа дня обнаружение флуоресценции может быть оптимизировано для сбора, поскольку предполагается, что источник сигнала (точечный) является фокусом падающего луча. Эта оптимизация сбора в сочетании с возбуждением в ближнем инфракрасном диапазоне приводит к общему преимуществу 2PM по сравнению с LSCM для получения изображений с большей глубины в толстых биологических образцах (например, MHB по всей оси DV).
Каждое событие поглощения, будь то линейное или нелинейное, потенциально катастрофично для эмиссионных свойств флуорофора. Фотообесцвечиванию способствует поглощенная энергия, благодаря которой фотоиндуцированное повреждение, химическая модификация и факторы окружающей среды способствуют выцветанию флуорофора. Поскольку поглощение энергии фотонов необходимо для процесса флуоресценции, возможность фотообесцвечивания неизбежна. В этом отношении 2PM более экономно, чем LSCM, в использовании флуорофоров, что может быть важным соображением при исследованиях изображений живых клеток в периоды развития (например,g., чтобы визуализировать развитие границы рестрикции линии MHB).
Недавние достижения в визуализации живых клеток привели к разработке систем, которые могут всесторонне фиксировать морфогенетические движения и деления на нескольких стадиях развития. Этим недавним достижениям в визуализации «in toto» способствовала способность сенсибилизировать методы микроскопии высокого разрешения к единичным ядрам с использованием генетически кодируемых флуоресцентных маркеров, таких как слияние гистона и зеленого флуоресцентного белка, хотя для визуализации и отслеживания использовались эндогенные сигналы. каждая клетка в эмбрионе рыбок данио для создания дерева клонов через первые 9 клеточных делений (Olivier et al., 2010). Локализованные в ядрах, флуоресцентные маркеры соседних клеток хорошо разделены, что важно для их выделения в тесноте эмбриона. Отслеживание клонов в этом контексте затем становится упражнением по отслеживанию предков и их потомков, хотя и сложным упражнением в области информатики, особенно в тканях с высокой плотностью клеток, таких как формирующийся MHB. Эта проблема способствовала развитию технологий визуализации и компьютерного анализа (Peng, 2008), которые выявили с высоким пространственным и временным разрешением коллективные миграции клеток (McMahon et al., 2008) и даже деления и движения каждой клетки в развивающемся эмбрионе рыбок данио в течение 24 часов (Keller et al., 2008).
Световая пластинчатая микроскопия, которая была разработана с использованием флуоресценции от линейного и нелинейного поглощения, недавно появилась в сообществе специалистов по биологии развития (Keller et al., 2010; Santi, 2011; Weber and Huisken, 2011; Huisken, 2012). несколько названий, включая SPIM (селективная микроскопия с плоским освещением), mSPIM (m для разнонаправленного) и DSLM (цифровая сканированная лазерная световая микроскопия), обозначающие различия в конфигурации и формировании светового листа (Keller and Stelzer, 2008).Базовый принцип SPIM был разработан в 1903 году Siedentopf и Zsigmondy (Huisken, 2012), но, как и в случае с LSCM, этот метод не оказал влияния на биологическое сообщество до тех пор, пока в 2004 году не использовался SPIM in vivo для изображения обоих относительно прозрачный эмбрион medaka и более непрозрачный эмбрион Drosophila (Huisken et al., 2004). Чтобы создать световой луч, Huisken et al. (2004) использовали цилиндрическую линзу, которая фокусирует свет вдоль одной оси вместо двух, как это делает сферическая линза, создавая световой слой, а не линию.Лист сканируется через образец, и сигнал обнаруживается линзой объектива, расположенной под углом 90 °, который отображает матрицу устройств с заряженной связью (ПЗС). Это параллельное возбуждение и обнаружение передает все изображение на камеру CCD, что значительно увеличивает скорость сбора данных, особенно по сравнению с точечной сканирующей микроскопией.
Совсем недавно световые листы были созданы путем быстрого сканирования лазерного луча с большой глубиной резкости или конфокальным параметром вдоль одной оси (DSLM), что обеспечивает значительно более высокое отношение сигнал / шум, чем предыдущие подходы (Keller and Stelzer, 2008).При демонстрации DSLM Keller and Stelzer (2008) охарактеризовали ядерные движения у рыбок данио в течение первых 24 часов развития как для эмбрионов дикого типа, так и для мутантных Mzoep. Keller и Stelzer (2008) обнаружили, что механизм образования гипобластов во время эпиболии варьирует в зависимости от положения, при этом дорсальная мезендодерма формируется за счет проникновения, а вентральная мезендодерма за счет инволюции. Мутанты Mzoep не смогли интернализовать клетки, чтобы сформировать гипобласт.
В конфигурации светового листа путь луча возбуждения находится в плоскости объекта обнаружения, и, таким образом, генерация сигнала флуоресценции, используемого для визуализации изображений, ограничивается плоскостью формирования изображения.Следовательно, независимо от того, вызвана ли флуоресценция линейным или нелинейным поглощением, возбужденные флуорофоры вносят свой вклад в рендеринг изображения до его неизбежной потери из-за фотообесцвечивания. Такое экономичное использование флуорофоров выгодно отличается от LSCM, который генерирует широкий сигнал и затем распознает расфокусированную флуоресценцию. Для оптимизации световой микроскопии для визуализации живых клеток и минимизации фотообесцвечивания, в частности, были разработаны различные конфигурации для максимизации скорости сбора данных и разрешения при гомогенизации освещения и минимизации пиковых интенсивностей, например.ж., решеточная световая микроскопия (Chen et al., 2014).
Теперь можно получать изображения с высоким (субклеточным) разрешением и отслеживать деления и морфогенетические движения каждой клетки в развивающемся эмбрионе на нескольких стадиях развития. Благодаря этим технологическим разработкам, а также развитию генетических и эмбриологических манипуляций для создания генетически закодированных молекулярных меток, теперь стало возможным визуализация отдельных компонентов и характеристика взаимодействий нескольких компонентов в развивающемся эмбрионе, что может сделать возможным многокомпонентный анализ сложных биологических систем.При использовании методов флуоресцентной микроскопии для многокомпонентной визуализации потребуются различные метки, возбуждение этих меток, их обнаружение и сегментация изображения.
Технические препятствия для многокомпонентной визуализации существуют, но быстро решаются. Одна из проблем — возбуждение и обнаружение нескольких флуорофоров в одном образце. Максимум излучения флуоресценции от линейного поглощения имеет меньшую энергию, чем поглощенный фотон, и, следовательно, спектрально смещен от длины волны его возбуждения.Таким образом, обнаружение сигнала флуоресценции может быть достигнуто с помощью спектрального отклонения возбуждающего лазера от тракта обнаружения. При использовании нескольких флуорофоров можно настроить возбуждение в пределах перекрывающихся спектров поглощения флуорофоров, настроить лазер возбуждения для каждого флуорофора или их комбинацию. Однако при использовании нескольких флуорофоров одна или несколько длин волн возбуждающего лазера могут перекрываться с сигналом флуоресценции. Следовательно, при спектральном отклонении возбуждающих лазеров часть сигнала также может быть отклонена.Один из способов избежать отклонения сигнала состоит в последовательном возбуждении флуорофоров, что также помогает в сегментации изображения, хотя и за счет скорости получения изображения (Valm et al., 2017).
Максимум излучения двухфотонно возбужденной флуоресценции выше по энергии, чем поглощенные фотоны. Фактически, длина волны возбуждения обычно спектрально далеко от окна обнаружения, то есть длины волн возбуждения в ближнем инфракрасном диапазоне для флуоресценции в видимой области. Таким образом, для возбуждения нескольких флуорофоров центральная длина волны возбуждающих лазерных импульсов может быть настроена в пределах перекрывающихся спектров нелинейного поглощения флуорофоров, можно настроить возбуждающий лазерный генератор для каждого флуорофора (хотя это не является рентабельным) или их комбинацию.Тем не менее, одновременное возбуждение и обнаружение нескольких флуорофоров возможно без потери сигнала из-за отклонения возбуждающих лазерных импульсов. Один из подходов заключается в использовании ультракороткого лазерного импульса с широким спектром возбуждения для нелинейного возбуждения нескольких флуорофоров одновременно (Gibbs et al., 2014a). Сегментация изображений может быть достигнута за счет спектрального разделения. Этот подход имеет дополнительное преимущество совместной регистрации изображений, поскольку для получения многокомпонентных изображений используется один лазер.В других схемах, например, элегантно использовалась нелинейная оптика для генерации многоцветных изображений формирования нейронных цепей в сочетании с системой маркировки «мозговой дуги» (Mahou et al., 2012).
Быстрый прогресс в технологиях визуализации в сочетании с новыми методами хромосомной инженерии с помощью CRISPR / Cas9 (например, см. Sander and Joung, 2014) обещает открыть новый период быстрых достижений, основанных на этих технологиях, чтобы понять развитие MHB. Например, способность конструировать определенные геномные локусы для экспрессии флуоресцентных репортеров важных для развития генов позволит лучше понять поведение клеточных клонов в организмах, удобных для визуализации.В качестве одного из примеров Ota et al. (2016) использовали нацеливание CRISPR / Cas для создания аллеля, экспрессирующего eGFP, у рыбок данио pax2a (Ota et al., 2016). Комбинируя репортеры клонов и цитоскелета в экспериментах по многокомпонентной визуализации, можно отслеживать вклад определенных типов клеток в морфогенез MHB. В качестве альтернативы, новые методы маркировки и визуализации с высоким разрешением могут позволить визуализировать определенные хромосомные локусы и энхансеры в определенных клеточных линиях, чтобы понять геномный контроль судьбы клеток внутри MHB.Эти новые инструменты быстро расширяют нашу способность визуализировать аспекты разработки MHB, которые ранее были скрыты от глаз.
Анализ изображений
Еще одним текущим ограничением для характеристики динамических изменений в развивающемся MHB является состояние программного обеспечения для анализа изображений. Количество и диапазон инструментов анализа изображений увеличивается. Использование этих инструментов варьируется от общих прикладных программ, таких как ImageJ и Icy (de Chaumont et al., 2012; Schindelin et al., 2015), до инструментов для конкретных приложений (Peng, 2008; Eliceiri et al., 2012). Развитие этих инструментов обусловлено необходимостью быстрого и количественного анализа больших мультимодальных наборов данных изображений. Такие данные различаются по типам изображений, временным рядам 2D или 3D изображений и многоканальным измерениям. Существует также множество аспектов анализа изображений, таких как визуализация пиксельных данных более информативными способами, выполнение автоматических измерений, когда изображения имеют отличительные особенности или ориентиры, и построение структурных моделей отображаемой системы. Несмотря на обилие доступного программного обеспечения, визуализация морфогенеза рыбок данио, особенно на ранних стадиях развития, представляет собой значительную проблему, поскольку различные ориентиры (Mikut et al., 2013), возможно, еще не развиты в этих системах. В настоящее время не существует единого программного обеспечения, которое могло бы обрабатывать стек изображений и визуализировать модель трехмерной реконструкции объема изображения. Примечательно, что даже несмотря на то, что эмбрион рыбок данио на ранней стадии относительно безликий (без ориентиров), люди могут легко распознать его форму. Тот факт, что человеческий мозг может обрабатывать изображение и строить мысленную карту эмбриона рыбок данио, предполагает, что в принципе возможно создание аналогичной 3D-модели с помощью вычислений.3D-модель относится к поверхностям и анатомическим объемам, «распознаваемым» или «идентифицированным» программным обеспечением и математически описываемым так, что возможна многомерная параметризация. Это относится к получению значений для 2- и 3-мерных характеристик, таких как измерение объема желудочков и отделов головного мозга, распределение экспрессии генов и пространственно-временная эволюция, а также совместная регистрация в разных образцах. Для достижения этой цели мы разработали программу под названием CAFE (Computer Aided Feature Extraction).Он возник из приложения, в основном для количественного определения волокон на 2D-изображениях (Hwang and Eryilmaz, 2014). Отличительной особенностью CAFE является его синергия между анализом изображений и построением моделей. Поскольку на пиксельные данные влияют экспериментальные условия, CAFE изначально использует пиксельные данные для построения крупнозернистой модели системы, отображаемой в виде шариков и палочек (рис. 6). Работа с геометрическими элементами вместо пиксельных данных увеличивает скорость вычислений и позволяет математическое описание модели. Например, для шаровой модели мозга рыбок данио другая модель из другого канала визуализации (например,g., стеки флуоресцентных изображений экспрессии генов) могут быть наложены друг на друга, что будет полезно для анализа пространственного отношения флуорофоров (рисунки 6A – C). После построения шаровой модели становятся возможными дополнительные математические операции, такие как определение нормалей к поверхности (рисунки 6D, E).
Рисунок 6 . Компьютерное извлечение признаков (CAFE) реконструкция изображений эмбрионов мозга рыбок данио. (A) Односрезовые изображения эмбриона мозга рыбок данио с двухфотонным микроскопом через 20 часов после оплодотворения, показывающие каналы автофлуоресценции (слева) и GFP (справа).Канал GFP отмечает wnt1 выражения. (B) Представление структуры эмбриона рыбок данио автофлуоресценции (проволочки) и GFP (зеленый объем). Визуализация изображений 1–45 в стеке из 79 изображений в дорсальной (слева) и вентральной (справа) проекциях. Автофлуоресценция (проволоки), реконструированная с использованием функции обнаружения на основе границ, для распознавания локальных градиентов пикселей и определения модели шариков и палочек по краям изображения. GFP (зеленый объем), реконструированный с использованием функции обнаружения на основе площади для размещения шариков в областях с высокой интенсивностью пикселей. (C) Детальная шариковая модель области в рамке в (B) , показывающая морфологическую поверхность, определенную CAFE (голубой цвет). (D) Локальные нормали поверхностей для поверхностей, выделенных в (C) . (E) Вид сверху наложения автофлуоресценции и реконструкций GFP с нормалями поверхности.
Для получения всеобъемлющего пространственно-временного атласа эмбрионального развития незаменим такой инструмент, как CAFE. В конечном итоге потребуется количественное описание процесса разработки.Например, эволюция объема и морфологии различных отделов мозга в различные моменты времени, а также распределение генов-репортеров по мозгу и их временная эволюция являются обязательными взаимосвязями, которые будут варьироваться в зависимости от патологического развития, включая заболевания нервной системы. Кроме того, чтобы построить «каноническую» модель развития мозга рыбок данио, измерения, описанные выше, должны быть выполнены на изображениях многих различных эмбрионов и усреднены в каждый момент времени, что требует совместной регистрации (Gibbs et al., 2014б). Построение таких канонических моделей является основной целью информатики структурных биоизображений.
Анализ между выборками данных позволит количественно оценить вариабельность морфогенетических изменений и предоставить определенную основу для выводов в исследованиях развития. Текущие инструменты обработки изображений различаются с точки зрения совместимости с модальностями визуализации, пользовательским интерфейсом и степенью автоматизированного анализа. Поскольку рыбки данио — идеальный модельный организм, который можно использовать для классификации как экспрессии генов, так и морфологии у позвоночных, существует несколько программных приложений, разработанных исключительно для высокопроизводительных данных визуализации рыбок данио.Применения варьируются от отслеживания клеток до создания трехмерного атласа мозга личинок рыбок данио, что позволяет исследователям решать вопросы биологии развития, нейронных путей, генетики и т. Д. (Ronneberger et al., 2012; Mikut et al., 2013; Gibbs et al. ., 2014б). Усовершенствованное приложение для рыбок данио — это ViBE-Z, веб-структура, которая выполняет совместную локализацию личинок рыбок данио через 48, 72 и 96 часов после оплодотворения (Ronneberger et al., 2012). ViBE-Z требует ручного выбора морфологических ориентиров из обучающего набора изображений для выполнения методов последовательной регистрации и согласования тестовых выборок с обучающим набором.Используя совместную локализацию изображений, ViBE-Z может сегментировать анатомические области личинки рыбок данио и визуализировать паттерны экспрессии генов в объеме рыбок данио. Ограничение на точки отбора проб, которые могут быть проанализированы, а также необходимое сходство между образцами для совместной локализации, создают проблему при анализе эмбрионов рыбок данио на ранней стадии (менее 24 часов оплодотворения) с минимальными основными характеристиками или мутантными фенотипами. Кроме того, основным ограничением ViBE-Z и другого программного обеспечения для визуализации рыбок данио является отсутствие возможностей для количественных измерений.Получение измерений на основе объема является сложной задачей, поскольку необходимы несколько изображений, эталонный набор координат и точные маркеры в определенном месте в пределах набора координат. Например, ViBE-Z соответствующим образом совместно регистрирует несколько образцов изображений и визуализирует, где, по отношению к анатомическим областям, ген выражается в объеме, но он не измеряет объем анатомических областей или распределение экспрессии генов, а также не учитывает временные сравнения.
CAFE удовлетворяет потребность в программных инструментах для анализа изображений, создавая объемную модель из стеков изображений на основе декартовой системы координат.CAFE использует реконструированную модель шарика и палки, основанную на локальной интенсивности пикселей исходного изображения, для выполнения этапов обработки изображения, таких как увеличение интенсивности или поворот изображения, а также для получения измерений 2D и 3D функций. Нормали к поверхности можно рассчитать по участкам поверхности, показанным на рисунке 6C. На рисунках 6D, E показаны эти нормали к поверхности. Для объемного анализа CAFE может обнаруживать границу, окружающую определяемую пользователем открытую область, и определять ее общую геометрию. Этот параметр является трехмерным дополнением параметра площади желудочка среднего мозга, полученного из двумерных поперечных сечений (Гиббс и др., 2014б). Алгоритмы на основе пикселей были разработаны и реализованы на нескольких платформах обработки изображений для выполнения расчетов анализа изображений. Способность CAFE создавать двумерные и трехмерные модели шариков и палочек и использовать эти представления для получения количественных расчетов расширяет возможности извлечения измерений из нескольких образцов изображений. Исследователи смогут количественно классифицировать пространственно-временные изменения в развивающемся эмбрионе и аналогичным образом отметить характерные эффекты отсутствующих генов-репортеров, например, в фенотипе ace .
Заключение
Важно понять его роль в развитии мозга и потенциальный вклад в формирование паттерна человеческого мозга врожденных дефектов. MHB также является замечательной моделью для разделения сигнального контроля судьбы клеток и морфогенеза тканей. В этом обзоре мы представляем доказательства того, что интерфейс передачи сигналов Wnt-Fgf коррелирует со специфическими морфогенетическими изменениями, которые управляют морфогенезом MHB. Сложность формирования паттерна и морфогенеза MHB создает императив для применения новых методологий и подходов, чтобы раскрыть лежащую в основе молекулярную природу.Одновременное создание правильной морфологии мозга, типов клеток и нейронных схем — сложная задача, но это очень надежный и адаптируемый процесс. Поведение клеток, такое как дифференциальная адгезия, рост и апоптоз, миграция и ремоделирование цитоскелета, должно быть точно скоординировано на больших участках внутри развивающегося нейроэпителия, и это достижение настолько поразительно, что понятно, что мы были одновременно очарованы и разочарованы нашими попытками понять этот процесс. в целом.Для продвижения к более полному механистическому пониманию, связывающему самые ранние события формирования паттернов с возможной архитектурой мозга и судьбами клеток, потребуется усиленное сотрудничество между дисциплинами, чтобы можно было сформулировать и тщательно протестировать наилучшие возможные модели. Есть еще много открытых вопросов относительно того, как формируется правильная архитектура мозга и какие типы клеток определяются во время развития нервной системы, хотя полное понимание этих процессов чрезвычайно важно с точки зрения фундаментальной науки, а также для развития регенеративной медицины при невральных заболеваниях.
Авторские взносы
Все авторы разработали и написали рукопись.
Финансирование
Работа поддержана Национальными институтами здравоохранения (грант R01NS088564). AC-G финансировалась стипендиями для аспирантов Национального научного фонда в рамках гранта № DGE 1252521. Любые мнения, выводы, выводы или рекомендации, выраженные в этом материале, принадлежат авторам и не обязательно отражают точку зрения Национального научного фонда. .
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы благодарят Дэвида Грина, Саурав Моханти и Эми Уайтенер за комментарии к рукописи. Некоторая информация, включенная в эту рукопись, была ранее опубликована в докторской диссертации Гиббса (2014), доступной в Интернете.
Список литературы
Бэбб С.Г., Барнетт Дж., Доеденс А.Л., Кобб Н., Лю К., Соркин Б.С. и др. (2001). E-кадгерин рыбок данио: экспрессия во время раннего эмбриогенеза и регуляция во время развития мозга. Dev. Дин. 221, 231–237. DOI: 10.1002 / dvdy.1132
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бэ, Ю. К., Кани, С., Симидзу, Т., Танабе, К., Нодзима, Х., Кимура, Ю. и др. (2009). Анатомия мозжечка рыбок данио и скрининг мутаций, влияющих на его развитие. Dev. Биол. 330, 406–426. DOI: 10.1016 / j.ydbio.2009.04.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пряжки, Г. Р., Торп, К. Дж., Рамель, М. К., и Леквен, А. С. (2004). Комбинаторный Wnt контроль образования границ между средним мозгом и задним мозгом рыбок данио. мех. Dev. 121, 437–447. DOI: 10.1016 / s0925-4773 (04) 00074-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чапутон, П., Адольф, Б., Лойхт, К., Тангейзер, Б., Рю, С., Дривер, В. и др. (2006). Экспрессия her5 выявляет пул нервных стволовых клеток в среднем мозге взрослых рыбок данио. Девелопмент 133, 4293–4303. DOI: 10.1242 / dev.02573
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Chen, B.C., Legant, W. R., Wang, K., Shao, L., Milkie, D. E., Davidson, M. W., et al. (2014). Решеточная световая микроскопия: визуализация молекул эмбрионов с высоким пространственно-временным разрешением. Наука 346: 1257998.DOI: 10.1126 / science.1257998
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чи, К. Л., Мартинес, С., Вурст, В., и Мартин, Г. Р. (2003). Сигнал истмического организатора FGF8 необходим для выживания клеток в предполагаемом среднем мозге и мозжечке. Девелопмент 130, 2633–2644. DOI: 10.1242 / dev.00487
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коэн, Э. Д., Тиан, Ю., и Морриси, Э. Э. (2008). Передача сигналов Wnt: важный регулятор сердечно-сосудистой дифференцировки, морфогенеза и самообновления предшественников. Девелопмент 135, 789–798. DOI: 10.1242 / dev.016865
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Купер С. Р., Эмонд М. Р., Дай П. К., Либау Б. Г., Вулман М. А. и Йонтес Дж. Д. (2015). Протокадгерины контролируют модульную сборку нейронных столбцов в покровной оболочке зрительного нерва рыбок данио. J. Cell Biol. 211, 807–814. DOI: 10.1083 / jcb.201507108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кокс, W.Г., и Хеммати-Бриванлу, А. (1995). Каудализация нервной судьбы за счет тканевой рекомбинации и bFGF. Развитие 121, 4349–4358.
PubMed Аннотация | Google Scholar
Дейли Л., Амброзетти Д., Мансухани А. и Базилико К. (2005). Механизмы, лежащие в основе дифференциальных ответов на передачу сигналов FGF. Cytokine Growth Factor Rev. 16, 233–247. DOI: 10.1016 / j.cytogfr.2005.01.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
де Шомон, Ф., Dallongeville, S., Chenouard, N., Herve, N., Pop, S., Provoost, T., et al. (2012). Icy: открытая платформа информатики биоизображений для расширенных воспроизводимых исследований. Нац. Методы 9, 690–696. DOI: 10.1038 / Nmeth.2075
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Доэрти Д., Миллен К. Дж., Баркович А. Дж. (2013). Пороки развития среднего и заднего мозга: достижения в области клинической диагностики, визуализации и генетики. Ланцет нейрол. 12, 381–393.DOI: 10,1016 / s1474-4422 (13) 70024-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Dyer, C., Blanc, E., Hanisch, A., Roehl, H., Otto, G.W., Yu, T., et al. (2014). Бимодальная функция передачи сигналов Wnt направляет градиент активности FGF, чтобы пространственно регулировать дифференцировку нейронов в среднем мозге. Развитие 141, 63–72. DOI: 10.1242 / dev.099507
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Элисейри, К. В., Бертольд, М.Р., Гольдберг, И. Г., Ибаньес, Л., Манджунат, Б. С., Мартоне, М. Э. и др. (2012). Программные средства биологической визуализации. Нац. Методы 9, 697–710. DOI: 10.1038 / Nmeth.2084
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Филас, Б.А., Олтеан, А., Маджиди, С., Бейли, П.В., Биби, Д.С., и Табер, Л.А. (2012). Региональные различия в сокращении актомиозина формируют первичные пузырьки в эмбриональном курином мозге. Phys. Биол. 9: 066007. DOI: 10.1088 / 1478-3975 / 9/6/066007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фуше, И., Миона, М., Симеоне, А., Акампора, Д., Балли-Куиф, Л., и Хоуарт, К. (2006). Дифференциация идентичностей клеток мозжечка в отсутствие передачи сигналов Fgf у морфантов Otx рыбок данио. Девелопмент 133, 1891–1900. DOI: 10.1242 / dev.02352
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гао, Л., Шао, Л., Хиггинс, К. Д., Поултон, Дж. С., Пайфер М., Дэвидсон М. В. и др. (2012). Неинвазивная визуализация за дифракционным пределом трехмерной динамики в сильно флуоресцентных образцах. Cell 151, 1370–1385. DOI: 10.1016 / j.cell.2012.10.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гато А. и Десмонд М. Э. (2009). Почему эмбрион все еще имеет значение: спинномозговая жидкость и нейроэпителий как взаимозависимые регуляторы роста, морфогенеза и гистиогенеза эмбрионального мозга. Dev. Биол. 327, 263–272.DOI: 10.1016 / j.ydbio.2008.12.029
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гиббс, Х. К. (2014). Оцифровка развития мозга позвоночных с помощью микроскопии ультракоротких импульсов. Техасский университет A&M: диссертация колледжа (Техас).
Google Scholar
Гиббс, Х.С., Бай, Ю.К., Леквен, А.С., и Йе, А.Т. (2014a). Визуализация эмбрионального развития с помощью микроскопии с ультракороткими импульсами. Опт. Англ. 53: 051506. DOI: 10.1117 / 1.oe.53.5.051506
CrossRef Полный текст | Google Scholar
Гиббс, Х.С., Додсон, К.Р., Бай, Ю.К., Леквен, А.С., и Йе, А.Т. (2014b). Комбинированное картирование клонов и профили экспрессии генов формирования паттерна мозга эмбриона с использованием микроскопии с ультракороткими импульсами и регистрации изображений. J. Biomed. Опт. 19: 126016. DOI: 10.1117 / 1.jbo.19.12.126016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гиббс, Х. К., Додсон, К. Р., Бай, Ю., Леквен, А.С., и Йе, А.Т. (2013). «Комбинированное картирование клонов и профилирование спецификаций судьбы с помощью NLOM-OCM с использованием импульсов длительностью менее 10 фс», в Proceedings SPIE 8593, Оптические методы в биологии развития . DOI: 10.1117 / 12.2004330
CrossRef Полный текст | Google Scholar
Грин Д., Уайтнер А. Э., Моханти С. и Леквен А. С. (2015). Постериоризация нервной системы позвоночных: оценка функции передачи сигналов wnt. Dev. Дин. 244, 507–512. DOI: 10.1002 / dvdy.24230
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гуцман Дж. Х., Грэден Э. Г., Лоури Л. А., Холли Х. С. и Сиве Х. (2008). Формирование сужения границы среднего и заднего мозга у рыбок данио требует ламинин-зависимого базального сужения. мех. Dev. 125, 974–983. DOI: 10.1016 / j.mod.2008.07.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гуцман, Дж. Х., Саху, С. У. и Квас, К. (2015).Немышечные миозины IIA и IIB по-разному регулируют изменения формы клеток во время морфогенеза мозга рыбок данио. Dev. Биол. 397, 103–115. DOI: 10.1016 / j.ydbio.2014.10.017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гуцман Дж. Х., Сиве Х. (2010). Эпителиальная релаксация, обеспечиваемая регулятором миозинфосфатазы Mypt1, необходима для расширения просвета желудочков мозга и морфогенеза заднего мозга. Развитие 137, 795–804. DOI: 10.1242 / дев.042705
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хардинг М. Дж., Нечипорук А. В. (2012). Передача сигналов Fgfr-Ras-MAPK необходима для апикального сужения посредством апикального позиционирования Rho-ассоциированной киназы во время формирования механосенсорных органов. Девелопмент 139, 3130–3135. DOI: 10.1242 / dev.082271
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Heimbucher, T., Murko, C., Bajoghli, B., Aghaallaei, N., Huber, A., Stebegg, R., et al. (2007). Gbx2 и Otx2 взаимодействуют с доменом WD40 корепрессоров Groucho / Tle. Мол. Клетка. Биол. 27, 340–351. DOI: 10.1128 / mcb.00811-06
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гейзенберг, К. П., Тада, М., Раух, Г. Дж., Сауде, Л., Конча, М. Л., Гейслер, Р. и др. (2000). Silberblick / Wnt11 обеспечивает конвергентные движения разгибания во время гаструляции рыбок данио. Природа 405, 76–81. DOI: 10.1038 / 35011068
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хойбергер, Дж., и Birchmeier, W. (2010). Взаимодействие опосредованной кадгерином клеточной адгезии и канонической передачи сигналов wnt. Колд Спринг Харб. Перспектива. Биол. 2: a002915. DOI: 10.1101 / cshperspect.a002915
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Идальго-Санчес, М., Миллет, С., Блох-Гальего, Э., и Альварадо-Малларт, Р. М. (2005). Спецификация мезо-истмо-мозжечковой области: граница Otx2 / Gbx2. Brain Res. Ред. 49, 134–149. DOI: 10.1016 / j.brainresrev.2005.01.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хьюскен, Дж., Свогер, Дж., Дель Бене, Ф., Виттбродт, Дж., И Стельцер, Э. Х. К. (2004). Оптическое срезы глубоко внутри живых эмбрионов с помощью микроскопии с селективным плоским освещением. Наука 305, 1007–1009. DOI: 10.1126 / science.1100035
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hwang, W., and Eryilmaz, E. (2014). Кинетическая подпись фрактально-подобных сетей нитей, образованных ориентационной линейной эпитаксией. Phys. Rev. Lett. 113: 025502. DOI: 10.1103 / Physrevlett.113.025502
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Иноуэ, Ф., Курокава, Д., Такахаши, М., и Аидзава, С. (2012). Gbx2 напрямую ограничивает экспрессию Otx2 передним и средним мозгом, конкурируя с факторами POU класса III. Мол. Клетка. Биол. 32, 2618–2627. DOI: 10.1128 / MCB.00083-12
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Jászai, J., Reifers, F., Пикер А., Лангенберг Т. и Бранд М. (2003). Трансформация перешейка в средний мозг при отсутствии активности организатора среднего и заднего мозга. Девелопмент 130, 6611–6623. DOI: 10.1242 / dev.00899
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Юнгблут, С., Ларсен, К., Визенманн, А., и Ламсден, А. (2001). Перемешивание клеток между эмбриональным средним мозгом и задним мозгом. Curr. Биол. 11, 204–207. DOI: 10,1016 / s0960-9822 (01) 00049-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Каша, К.Э., Заллен Дж. А. (2011). Динамика и регуляция сократительных актин-миозиновых сетей в морфогенезе. Curr. Opin. Cell Biol. 23, 30–38. DOI: 10.1016 / j.ceb.2010.10.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Келлер, П. Дж., Шмидт, А. Д., Сантелла, А., Хайри, К., Бао, З. Р., Виттбродт, Дж. И др. (2010). Быстрая высококонтрастная визуализация развития животных с помощью микроскопии со структурированным освещением на основе сканируемых световых пластин. Нац. Методы 7, 637–642.DOI: 10.1038 / nmeth.1476
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Келлер П., Шмидт А., Виттбродт Дж. И Стельцер Э. (2008). Реконструкция раннего эмбрионального развития рыбок данио с помощью сканирующей световой микроскопии. Наука 322, 1065–1069. DOI: 10.1126 / science.1162493
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Келлер, П. Дж., И Стельцер, Э. Х. К. (2008). Количественное изображение in vivo целых эмбрионов с помощью цифровой сканированной лазерной флуоресцентной микроскопии. Curr. Opin. Neurobiol. 18, 624–632. DOI: 10.1016 / j.conb.2009.03.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Киккер К. и Нирс К. (2001). Градиент морфогена передачи сигналов Wnt / beta-catenin регулирует формирование переднезаднего нейрального паттерна у Xenopus. Девелопмент 128, 4189–4201.
PubMed Аннотация | Google Scholar
Ким, С. А., Тай, К. Ю., Мок, Л. П., Моссер, Э. А., и Шуман, Э. М. (2011). Кальций-зависимая динамика взаимодействий кадгерина в межклеточных соединениях. Proc. Natl. Акад. Sci. U S A 108, 9857–9862. DOI: 10.1073 / pnas.10108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кишимото, Н., Альфаро-Сервелло, К., Симидзу, К., Асакава, К., Урасаки, А., Нонака, С., и др. (2011). Миграция предшественников нейронов из зоны конечного желудочка в обонятельную луковицу у взрослых рыбок данио. J. Comp. Neurol. 519, 3549–3565. DOI: 10.1002 / cne.22722
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Климковский, М.В., Росси, К. С., и Артингер, К. Б. (2010). Механизмы, управляющие индукцией и миграцией нервного гребня у рыбок данио и Xenopus laevis. Celll Adhes. Мигр. 4, 595–608. DOI: 10.4161 / cam.4.4.12962
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кносп, В. М., Нокс, С. М., Ломберт, И. М. А., Хэддокс, К. Л., Патель, В. Н., и Хоффман, М. П. (2015). Поднижнечелюстной парасимпатический ганглиогенез требует зависимых от ростков сигналов Wnt от эпителиальных предшественников. Dev. Cell 32, 667–677. DOI: 10.1016 / j.devcel.2015.01.023
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Курокава Д., Омура Т., Акасака К. и Айзава С. (2012). Клон-специфический энхансер управляет экспрессией Otx2 в тканях организатора костистых костей. мех. Dev. 128, 653–661. DOI: 10.1016 / j.mod.2011.11.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лангенберг, Т., и Бранд, М. (2005). Ограничение клонов поддерживает стабильную популяцию клеток-организаторов на границе среднего и заднего мозга рыбок данио. Девелопмент 132, 3209–3216. DOI: 10.1242 / dev.01862
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лапеби П., Боркиеллини К. и Хоулистон Э. (2011). Рассечение пути PCP: один или несколько путей? Управляет ли отдельный путь Wnt-Fz-Rho морфогенезом? Bioessays 33, 759–768. DOI: 10.1002 / bies.201100023
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ласситер Р. Н., Старк М. Р., Чжао Т.Ю., Чжоу, К. Дж. Дж. (2014). Сигнальные механизмы, контролирующие нейрогенез и расслоение краниальных плакод. Dev. Биол. 389, 39–49. DOI: 10.1016 / j.ydbio.2013.11.025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Леквен, А. К., Баклс, Г. Р., Костакис, Н., и Мун, Р. Т. (2003). wnt1 и wnt10b функционируют избыточно на границе среднего и заднего мозга рыбок данио. Dev. Биол. 254, 172–187. DOI: 10.1016 / s0012-1606 (02) 00044-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Леле, З., Folchert, A., Concha, M., Rauch, G.J., Geisler, R., Rosa, F., et al. (2002). Парашют / n-кадгерин необходим для морфогенеза и поддержания целостности нервной трубки рыбок данио. Развитие 129, 3281–3294.
PubMed Аннотация | Google Scholar
Лин, М. Дж., И Ли, С. Дж. (2016). Stathmin-like 4 критически важен для поддержания нейральных клеток-предшественников в дорсальной части среднего мозга личинок рыбок данио. Sci. Реп. 6: 36188. DOI: 10.1038 / srep36188
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лоури, Л.А., Де Риенцо, Г., Гуцман, Дж. Х., и Сиве, Х. (2009). Характеристика и классификация мутантов морфологии мозга рыбок данио. Анат. Рек. (Хобокен) 292, 94–106. DOI: 10.1002 / ar.20768
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лоури, Л. А., Сиве, Х. (2004). Стратегии нейруляции позвоночных и переоценка формирования нервной трубки костистой кости. мех. Dev. 121, 1189–1197. DOI: 10.1016 / j.mod.2004.04.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Махоу, П., Zimmerley, M., Loulier, K., Matho, K. S., Labroille, G., Morin, X., et al. (2012). Многоцветная двухфотонная визуализация тканей путем смешивания длин волн. Нац. Методы 9, 815–818. DOI: 10.1038 / Nmeth.2098
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мартинес С., Кроссли П. Х., Кобос И., Рубенштейн Дж. Л. Р. и Мартин Г. Р. (1999). FGF8 индуцирует образование эктопического истмического организатора и развитие перешейка мозжечка посредством репрессивного действия на экспрессию Otx2. Девелопмент 126, 1189–1200.
PubMed Аннотация | Google Scholar
МакГрю, Л. Л., Лай, К.-Дж., и Мун, Р. Т. (1995). Спецификация переднезадней нервной оси посредством синергетического взаимодействия сигнального каскада Wnt с noggin и фоллистатином . Dev. Биол. 172, 337–342. DOI: 10.1006 / dbio.1995.0027
PubMed Аннотация | CrossRef Полный текст | Google Scholar
МакМэхон, А.П., Брэдли, А.(1990). Протоонкоген Wnt-1 (int-1) необходим для развития большой области мозга мыши. Cell 62, 1073–1085. DOI: 10.1016 / 0092-8674 (90)
-рPubMed Аннотация | CrossRef Полный текст | Google Scholar
МакМахон А., Супатто В., Фрейзер С. Э. и Статопулос А. (2008). Динамический анализ гаструляции Drosophila дает представление о коллективной миграции клеток. Наука 322, 1546–1550. DOI: 10.1126 / science.1167094
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мегасон, С.Г. и МакМахон А. П. (2002). Градиент митогена дорсальной средней линии Wnts организует рост в ЦНС. Развитие 129, 2087–2098.
PubMed Аннотация | Google Scholar
Мейер, Ф., Гизерт, Ф., Делик, С., Фаус-Кесслер, Т., Матеус, Ф., Симеоне, А. и др. (2014). Передача сигналов FGF / FGFR2 регулирует образование и правильное положение глиальных клеток Бергмана в развивающемся мозжечке мыши. PLoS One 9: e101124. DOI: 10.1371 / journal.pone.0101124
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Микут, р., Dickmeis, T., Driever, W., Geurts, P., Hamprecht, F.A., Kausler, B.X., et al. (2013). Автоматическая обработка данных изображений рыбок данио: обзор. Данио 10, 401–421. DOI: 10.1089 / zeb.2013.0886
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Миллен К. Дж., Стешина Е. Ю., Искусных И. Ю., Чижиков В. В. (2014). Трансформация мозжечка в более вентральную часть ствола мозга вызывает агенез мозжечка в отсутствие функции ptf1a. Proc. Natl. Акад. Sci. U S A 111, E1777 – E1786. DOI: 10.1073 / pnas.1315024111
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мияке А., Нихно С., Муракоши Ю., Сацука А., Накаяма Ю. и Ито Н. (2012). Neucrin, новый секретируемый антагонист канонической передачи сигналов Wnt, играет роль в развитии нервных тканей у рыбок данио. мех. Dev. 128, 577–590. DOI: 10.1016 / j.mod.2012.01.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нинкович, Я., Tallafuss, A., Leucht, C., Topczewski, J., Tannhäuser, B., Solnica-Krezel, L., et al. (2005). Ингибирование нейрогенеза на границе среднего и заднего мозга рыбок данио за счет комбинированной и дозозависимой активности новой пары генов hairylE (spl). Развитие 132, 75–88. DOI: 10.1242 / dev.01525
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нюхольм, М. К., Абделила-Сейфрид, С., и Гринблат, Ю. (2009). Новый генетический механизм регулирует формирование дорсолатеральных шарниров во время черепной нейруляции у рыбок данио. J. Cell Sci. 122, 2137–2148. DOI: 10.1242 / jcs.043471
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Olivier, N., Luengo-Oroz, M. A., Duloquin, L., Faure, E., Savy, T., Veilleux, I., et al. (2010). Реконструкция клеточного клона ранних эмбрионов рыбок данио с использованием нелинейной микроскопии без метки. Наука 329, 967–971. DOI: 10.1126 / science.1189428
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ота, С., Таймацу, К., Янаги, К., Намики, Т., Ога, Р., Хигасидзима, С., и др. (2016). Функциональная визуализация и нарушение целевых генов с использованием CRISPR / Cas9-опосредованной интеграции репортера eGFP в zebrafiah. Sci. Реп. 6: 34991. DOI: 10.1038 / srep34991
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Парада К., Мартин К., Алонсо М. И., Моро Дж. А., Буэно Д. и Гато А. (2005). Эмбриональная спинномозговая жидкость взаимодействует с истмическим организатором, чтобы регулировать экспрессию мезэнцефальных генов. J. Neurosci. Res. 82, 333–345. DOI: 10.1002 / jnr.20618
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Паркер М. О., Брок А. Дж., Уолтон Р. Т. и Бреннан К. Х. (2013). Роль рыбок данио (Danio rerio) в анализе генетики и нейронных цепей исполнительной функции. Фронт. Нейронные схемы 7:13. DOI: 10.3389 / fncir.2013.00063
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пикер, А., Бреннан, К., Рейферс, Ф., Кларк, Дж. Д. У., Холдер, Н., и Бранд, М. (1999). Требование к границе среднего и заднего мозга у рыбок данио в поляризации среднего мозга, картировании и ограничении ретинотектальной проекции. Развитие 126, 2967–2978.
PubMed Аннотация | Google Scholar
Puelles, E., Acampora, D., Lacroix, E., Signore, M., Annino, A., Tuorto, F., et al. (2003). Otx дозозависимый интегрированный контроль переднезаднего и дорсо-вентрального паттерна среднего мозга. Нац.Neurosci. 6, 453–460. DOI: 10.1038 / nn1037
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Rapaciolii, M., Palma, V., and Flores, V. (2016). Морфогенетические и гистогенетические роли пространственно-временной организации клеточной пролиферации в кортикогенезе позвоночных, выявленные в результате межвидового анализа развития коры тектума зрительного нерва. Фронт. Клетка. Neurosci. 10: 112. DOI: 10.3389 / fncel.2016.00112
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Речер, Г., Jouralet, J., Brombin, A., Heuzé, A., Mugniery, E., Hermel, J.M, et al. (2013). Медленно амплифицирующиеся предшественники среднего мозга рыбок данио обнаруживают высокие уровни транскриптов для нуклеотидного и рибосомного биогенеза. Развитие 140, 4860–4869. DOI: 10.1242 / dev.099010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Райфер Ф., Бели Х., Уолш Э. К., Кроссли П. Х., Стейнир Д. Ю. Р. и Бранд М. (1998). Fgf8 мутирован у acerebellar (ace) мутантов рыбок данио и необходим для поддержания развития границ среднего и заднего мозга и сомитогенеза. Девелопмент 125, 2381–2395.
PubMed Аннотация | Google Scholar
Рейм Г. и Бранд М. (2002). Spiel-ohne-grenzen / pou2 обеспечивает региональную компетентность в ответ на Fgf8 во время раннего нервного развития рыбок данио. Развитие 129, 917–933.
PubMed Аннотация | Google Scholar
Ринн, М., Лун, К., Арендт, Р., Геффарт, М., и Бранд, М. (2009). Gbx1 рыбок данио уточняет границу среднего и заднего мозга и обеспечивает сигнал постериоризации Wnt8. Neural Dev. 4:12. DOI: 10.1186 / 1749-8104-4-12
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ринн, М., Лун, К., Аморес, А., Ян, Ю. Л., Постлетвейт, Дж. Х. и Бранд, М. (2003). Клонирование, экспрессия и связь генов gbx1 и gbx2 рыбок данио с передачей сигналов Fgf. мех. Dev. 120, 919–936. DOI: 10.1016 / s0925-4773 (03) 00135-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ринн, М., Лун, К., Луз, М., Вернер, М., и Бранд, М. (2005). Позиционирование пограничного организатора среднего и заднего мозга посредством глобальной постериоризации нейроэктодермы, опосредованной передачей сигналов Wnt8. Развитие 132, 1261–1272. DOI: 10.1242 / dev.01685
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роннебергер О., Лю К., Рат М., Рюсс Д., Мюллер Т., Скиббе Х. и др. (2012). ViBE-Z: платформа для трехмерного виртуального анализа колокализации в мозге личинок рыбок данио. Нац.Методы 9, 735–742. DOI: 10.1038 / nmeth.2076
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Саху, С. У., Висецук, М. Р., Гард, Р. Дж., Хеннес, Л., Квас, К., и Гуцман, Дж. Х. (2017). Сигналы кальция управляют изменением формы клеток во время формирования границы между средним мозгом и задним мозгом рыбок данио. Мол. Биол. Cell 28, 875–882. DOI: 10.1091 / mbc.E16-08-0561
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шинделин, Дж., Рюден, К.Т., Хайнер, М.С., и Элисейри, К.В. (2015). Экосистема ImageJ: открытая платформа для анализа биомедицинских изображений. Мол. Репродукция. Dev. 82, 518–529. DOI: 10.1002 / mrd.22489
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шмитц Б., Папан К. и Кампос-Ортега Дж. А. (1993). Невруляция в переднем отделе туловища у рыбок данио brachydanio-rerio. Rouxs Arch. Dev. Биол. 202, 250–259. DOI: 10.1007 / BF00363214
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Северино, М., Аллегри, А. Э. М., Писторио, А., Ровильоне, Б., Ди Йорги, Н., Магни, М. и др. (2014). Вовлечение среднего и заднего мозга в септооптическую дисплазию. г. J. Neuroradiol. 35, 1586–1592. DOI: 10.3174 / ajnr.A3959
CrossRef Полный текст | Google Scholar
Смутный, М., Акос, З., Сильвия, Г., Шамипур, С., Рупрехт, В., Чапек, Д., и др. (2017). Силы трения позиционируют нервный зачаток. Нац. Cell Biol. 19, 306–317. DOI: 10.1038 / ncb3492
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Станганелло, Э., Хагеманн, А.И.Х., Маттес, Б., Синнер, К., Мейен, Д., Вебер, С. и др. (2015). Транспорт Wnt на основе филоподий во время формирования паттерна ткани позвоночных. Нац. Commun. 6: 5846. DOI: 10.1038 / ncomms6846
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сунмону, Н. А., Ли, К. Р., Го, К. X., и Ли, Дж. Ю. Х. (2011). Gbx2 и Fgf8 последовательно необходимы для формирования границы компартмента среднего и заднего мозга. Девелопмент 138, 725–734. DOI: 10.1242 / дев.055665
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Таллафус, А., и Балли-Куиф, Л. (2003). Отслеживание потомства her5 у трансгенных рыбок данио показывает динамику нейрогенеза и поддержания среднего и заднего мозга. Девелопмент 130, 4307–4323. DOI: 10.1242 / dev.00662
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Томас К. Р. и Капеччи М. Р. (1990). Целенаправленное разрушение протоонкогена int-1 мыши, приводящее к серьезным нарушениям в развитии среднего мозга и мозжечка. Природа 346, 847–850. DOI: 10.1038 / 346847a0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Tossell, K., Andreae, L.C, Cudmore, C., Lang, E., Muthukrishnan, U., Lumsden, A., et al. (2011a). Lrrn1 необходим для образования границы среднего и заднего мозга и организатора посредством регуляции различий сродства между клетками среднего и заднего мозга у цыплят. Dev. Биол. 352, 341–352. DOI: 10.1016 / j.ydbio.2011.02.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тосселл, К., Киккер, К., Визенманн, А., Ланг, Э., и Ирвинг, К. (2011b). Передача сигналов Notch стабилизирует формирование границ в организаторе среднего и заднего мозга. Девелопмент 138, 3745–3757. DOI: 10.1242 / dev.070318
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цай, Ф. К., Куо, Г. Х., Чанг, С. В., и Цай, П. Дж. (2015). Передача сигналов Ca 2+ при реорганизации цитоскелета, миграции клеток и метастазировании рака. Biomed Res. Int. 2015: 409245.DOI: 10.1155 / 2015/409245
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Туазон, Ф. Б., и Маллинс, М. С. (2015). Координированные во времени сигналы постепенно формируют переднезаднюю и дорсовентральную оси тела. Семин. Cell Dev. Биол. 42, 118–133. DOI: 10.1016 / j.semcdb.2015.06.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Валм А., Коэн С., Легант В., Мелунис Дж., Хершберг У., Уэйт Э. и др. (2017).Применение спектральной визуализации и анализа на системном уровне для выявления взаимодействия органелл. Природа 546, 162–167. DOI: 10.1038 / природа22369
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Varelas, X., Bouchie, M. P., and Kukuruzinska, M. A. (2014). N-гликозилирование белков при раке полости рта: нарушение регуляции клеточных сетей среди DPAGT1, адгезия E-кадгерина и каноническая передача сигналов Wnt. Гликобиология 24, 579–591. DOI: 10.1093 / glycob / cwu031
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фолькманн, К., Чен, Ю. Ю., Харрис, М. П., Вуллиманн, М. Ф., и Кёстер, Р. В. (2010). Верхняя ромбическая губа мозжечка рыбок данио генерирует тегментальные ядра заднего мозга за счет миграции на большие расстояния эволюционно консервативным способом. J. Comp. Neurol. 518, 2794–2817. DOI: 10.1002 / cne.22364
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ву К. и Фрейзер С. Э. (1995). Порядок и последовательность в карте судьбы нервной системы рыбок данио. Развитие 121, 2595–2609.
PubMed Аннотация | Google Scholar
Юк, Дж. И., Ли, X. Y., Ота, И., Фирон, Э. Р., и Вайс, С. Дж. (2005). Wnt-зависимая регуляция репрессора E-кадгерина улитки. J. Biol. Chem. 280, 11740–11748. DOI: 10.1074 / jbc.m413878200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Yu, S. R., Burkhardt, M., Nowak, M., Ries, J., Petrásek, Z., Scholpp, S., et al. (2009). Градиент морфогена Fgf8 формируется по механизму источник-сток со свободно диффундирующими молекулами. Природа 461, 533–536. DOI: 10.1038 / nature08391
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Зервас, М., Миллет, С., Ан, С., и Джойнер, А. Л. (2004). Поведение клеток и генетические линии среднего мозга и ромбомера 1. Neuron 43, 345–357. DOI: 10.1016 / j.neuron.2004.07.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Как работает Intersect — ArcGIS Pro | Документация
Инструмент Пересечение вычисляет геометрическое пересечение любого количества классов пространственных объектов и векторных слоев.Объекты или часть объектов, которые являются общими для всех входных данных (т. Е. Пересекаются), будут записаны в выходной класс пространственных объектов.
Intersect выполняет следующие функции:
- Определяет пространственную привязку для обработки. Это также будет пространственная привязка выходного класса пространственных объектов. Подробнее о том, как это делается, см. Пространственная привязка и геообработка. Все входные объекты проецируются в эту пространственную привязку для обработки.
- Трещины и скопления деталей.Крекинг вставляет вершины на пересечении краев элементов; кластеризация соединяет вместе вершины, которые находятся в пределах допуска xy.
- Обнаруживает геометрические взаимосвязи (пересечения) между объектами из всех классов пространственных объектов или слоев.
- Записывает эти пересечения как объекты (точки, линии или полигоны) на выходе.
Когда в списке входных объектов указано несколько классов пространственных объектов или слоев, порядок записей в списке не влияет на тип выходных объектов, но на пространственную привязку верхней записи в диалоговом окне инструмента (первая запись в скрипте) в списке будет использоваться во время обработки и устанавливаться для вывода.
Чтобы явно управлять выходной пространственной привязкой (системой координат и доменами), переопределите свойства пространственной привязки по умолчанию, используя соответствующие среды геообработки.
Входными данными может быть любая комбинация типов геометрии (точка, многоточечный, линейный или многоугольный). Тип выходной геометрии может иметь только ту же геометрию или геометрию меньшего размера, что и входной класс пространственных объектов с геометрией самого низкого измерения (точка = измерение 0, линия = 1 измерение и poly = 2 измерения).Указание другого типа выходных данных приведет к разным типам пересечений входных классов пространственных объектов. Это не разные представления одних и тех же перекрестков; они являются пересечениями, которые могут быть представлены только этим типом геометрии (точка, линия или многоугольник).
Совет:
Intersect может работать с одним входом. В этом случае вместо обнаружения пересечений между объектами из разных классов пространственных объектов или слоев он обнаружит пересечения между объектами в пределах одного входа.Это может быть полезно для обнаружения наложения полигонов и пересечений линий (в виде точек или линий).
Примеры: Входные полигоны
Полигоны могут пересекаться тремя способами:
- Перекрытие — область перекрытия можно создать, оставив для Типа вывода значение по умолчанию (НАИМЕНЬШЕЕ).
- Общая граница / касание на линии — этот тип пересечения можно создать, указав ЛИНИЮ в качестве Типа вывода.
- Касание в точке — этот тип пересечения можно создать, указав ТОЧКУ в качестве типа вывода.
Входные и выходные полигоны
На приведенном ниже рисунке показан результат пересечения двух классов полигональных объектов с параметром Тип вывода, установленным либо на POLY, либо на значение по умолчанию (LOWEST). Выходные полигональные объекты — это места, где полигон одного из входных классов пространственных объектов или слоя пересекает полигон другого входного класса пространственных объектов или слоя.
Входные и линейные полигоны
На рисунке ниже показан результат пересечения двух классов полигональных объектов с параметром Тип вывода, установленным на ЛИНИЯ.Выходные линейные объекты — это место, где полигон одного из входных классов пространственных объектов имеет общую границу (пересекается по линии) с полигоном из другого входного класса пространственных объектов.
Входные и точечные полигоны
На рисунке ниже показан результат пересечения двух классов полигональных объектов с параметром Тип вывода, установленным на ТОЧКА. Выходные точечные объекты — это места, где у полигона одного из входных классов пространственных объектов есть вершина, пересекающая границу (пересекающуюся в точке) полигона из другого входного класса пространственных объектов.
Пересечь полигон с точкойПримеры: линейные входные данные
Когда все входные данные являются классами линейных объектов, инструмент пересечения можно использовать для определения, где объекты из входных классов пространственных объектов перекрываются и пересекаются в точках и линиях.
Линейные входы и линейные выходы. На приведенном ниже рисунке показан результат пересечения двух классов линейных объектов с параметром Тип вывода, установленным на LOWEST или LINE. Выходные линейные объекты — это место, где линия одного из входных классов пространственных объектов перекрывает объект из другого входного класса пространственных объектов.
Линейные входы и точечные выходы
На рисунке ниже показан результат пересечения двух классов линейных объектов с параметром Тип вывода, установленным на ТОЧКА. Выходные точечные объекты — это место, где линия одного из входных классов пространственных объектов пересекает объект из другого входного класса пространственных объектов.
Пример: точечные входные данные
Если все входные данные являются классами точечных объектов, инструмент Пересечение можно использовать для определения точек, общих для всех входных классов пространственных объектов.
Примеры: входные данные смешанной геометрии
Пересечение можно использовать с классами пространственных объектов различной геометрии. Тип вывода по умолчанию (и максимально допустимый) такой же, как у класса пространственных объектов с геометрией наименьшего размера.
Полигон и линейный ввод для линейного вывода
На рисунке ниже показан результат пересечения классов линейных и полигональных объектов с параметром Тип вывода, установленным на ЛИНИЯ. Выходные линейные объекты — это место, где линия одного из входных классов пространственных объектов перекрывает полигон другого входного класса пространственных объектов.
Полигон и линейный вход для точечного вывода
На рисунке ниже показан результат пересечения классов линейных и полигональных объектов с параметром Тип вывода, установленным на ТОЧКА. Выходные точечные объекты — это места, где конечные точки линий касаются точки на границе полигона, а линии пересекают границы полигона. На выходе не создаются точки, если линии лежат непосредственно вдоль границ многоугольника.
Входные полигоны, линии и точки для выходных данных
На приведенном ниже рисунке показан результат пересечения классов пространственных объектов точек, линий и полигонов.Результатом может быть только класс точечных объектов. Каждая точка в выходных данных будет пересекать по крайней мере один объект в каждом из входных классов объектов.
Связанные темы
Отзыв по этой теме?
О методе нормального пересечения границ для генерации эффективного фронта
Abstract
В данной статье рассматривается проблема нахождения репрезентативной выборки оптимальных по Парето точек в многокритериальной оптимизации.Алгоритм нормального пересечения границ — это схема скаляризации для генерации набора равномерно распределенных эффективных решений. Недостатком этого алгоритма является то, что оптимальность решения по Парето не гарантируется. Вклад этой статьи двоякий. Во-первых, он представляет альтернативную формулировку этого алгоритма, гарантирующую (слабую) Парето-оптимальность решений. Это улучшение делает этот алгоритм теоретически эквивалентным другим классическим алгоритмам (таким как методы взвешенной суммы или ε -ограничений) без потери его способности генерировать набор равномерно распределенных эффективных решений.Во-вторых, представлен алгоритм, позволяющий заранее знать об определенных подзадачах, решения которых не являются оптимальными по Парето, и, таким образом, не тратить зря вычислительные усилия на их решение. Также представлена взаимосвязь нового алгоритма с методами взвешенной суммы и целевого программирования.
Ключевые слова
Многоцелевая оптимизация Эффективная генерация фронта Эффективный с вычислительной точки зрения алгоритм Скачатьпрочитать полный текст доклада конференции
Список литературы
1.
Дас, И., Деннис, Дж. Э .: Более пристальный взгляд на недостатки минимизации взвешенной суммы объектов для генерации множеств Парето в задачах многокритериальной оптимизации. Структурная оптимизация 14 (1), 63–69 (1997)
CrossRefGoogle Scholar2.
Дас, И., Деннис, Дж. Э .: Нормально-граничное пересечение: новый метод построения поверхности Парето в нелинейных задачах многокритериальной оптимизации . Журнал оптимизации SIAM 8 (3), 631–657 (1998)
zbMATHCrossRefMathSciNetGoogle Scholar3.
Деб, К .: Многоцелевая оптимизация с использованием эволюционных алгоритмов. Уайли, Чичестер (2001)
zbMATHGoogle Scholar4.
5.
Эшенауэр, Х., Коски, Дж., Осичка, А.: Оптимизация многокритериального проектирования. Springer, Berlin (1990)
zbMATHGoogle Scholar6.
Гембицки Ф.У .: Оптимизация производительности и чувствительности: метод векторного индекса. Докторская диссертация, Университет Кейс Вестерн Резерв, Кливленд, Огайо (1974)
Google Scholar7.
Ким, И.Ю., де Век, О.: Многокритериальная оптимизация с помощью метода адаптивной взвешенной суммы. В: 10-я Конференция по междисциплинарному анализу и оптимизации AIAA / ISSMO (2004)
Google Scholar8.
Ким, И.Я., де Век, О.: Метод адаптивной взвешенной суммы для многокритериальной оптимизации: новый метод генерации фронта Парето. Структурная и междисциплинарная оптимизация 31 (2), 105–116 (2005)
CrossRefGoogle Scholar9.
Messac, A., Маттсон, К.А.: Нормальный метод ограничения с гарантией равномерного представления полной границы Парето. AIAA Journal 42 (10), 2101–2111 (2004)
CrossRefGoogle Scholar10.
11.
Asyczka, A .: Эволюционные алгоритмы для оптимизации дизайна с одним и несколькими критериями. Physica Verlag, Heidelberg (2002)
zbMATHGoogle Scholar12.
Sandgren, E .: Оптимизация многокритериального проектирования с помощью целевого программирования. В: Адели, Х.(ред.) Достижения в оптимизации дизайна, стр. 225–265. Chapman & Hall, Лондон (1994)
Google Scholar13.
Стаников Р. Б., Матусов Дж. Б. Многокритериальная оптимизация и инженерия. Chapman and Hall, New York (1995)
Google Scholar
Информация об авторских правах
© Springer Berlin Heidelberg 2007
Авторы и филиалы
- 1. Институт вычислительной математики, факультет математики, Дрезденский технологический университет, Дрезден PIN 01069 Германия
О движении жидкости в пограничном слое вблизи линии пересечения двух плоскостей
Версия PDF также доступна для скачивания.
Кто
Люди и организации, связанные либо с созданием этого отчета, либо с его содержанием.
Какие
Описательная информация, помогающая идентифицировать этот отчет.Перейдите по ссылкам ниже, чтобы найти похожие предметы в Электронной библиотеке.
Когда
Даты и периоды времени, связанные с этим отчетом.
Статистика использования
Когда последний раз использовался этот отчет?
Взаимодействовать с этим отчетом
Вот несколько советов, что делать дальше.
Версия PDF также доступна для скачивания.
Ссылки, права, повторное использование
Международная структура взаимодействия изображений
Распечатать / Поделиться
Печать
Электронная почта
Твиттер
Facebook
Tumblr
Reddit
Ссылки для роботов
Полезные ссылки в машиночитаемых форматах.
Ключ архивных ресурсов (ARK)
Международная структура взаимодействия изображений (IIIF)
Форматы метаданных
Изображений
URL
Статистика
Лойцианский, Л.Г., Большаков, В. П. О движении жидкости в пограничном слое вблизи линии пересечения двух плоскостей. отчет, Ноябрь 1951 г .; (https://digital.library.unt.edu/ark:/67531/metadc62989/: по состоянию на 18 апреля 2021 г.), Библиотеки Университета Северного Техаса, Цифровая библиотека UNT, https://digital.library.unt.edu; кредитование Департамента государственных документов библиотек ЕНТ.
Подход к определению границ группы перекрестка для анализа трафика в связи с улучшением перекрестка
[1] Ягода, Х.N., et al. Разделение сигнальных систем на зоны управления. Транспортная инженерия, Vol. 43 No. 12, 1973, pp.42-46.
[2] Чин-Пинг, Э.Как принять решение о подключении изолированных дорожных сигналов. 1985. Нью-Йорк, Нью-Йорк, США: IEEE.
[3] Крюк, Д.и А. Альберс. Сравнение альтернативных методологий для определения точек останова в прохождении сигнала. на 69-м ежегодном собрании Института инженеров транспорта. 1999 г. Лас-Вегас, Невада.
[4] Хуш, Д.и Дж. Олбек. Руководство пользователя Synchro 6. Trafficware, Олбани, Калифорния, (2003).
[5] Гордон, Р.Л. и У. Тайге, Справочник по системам управления движением. 2005, FHWA: Вашингтон Д. К.
[6] Май., Ю. Цзэн и X. Ян. Исследование степени корреляции для проектирования движения группы перекрестков. Журнал Пекинского технологического института, Vol. 21 № Доп. 2, 2012, с. 11-14.
[7] Новый Южный Уэльс, Р.а. T. A. o., Руководство пользователя Scats. 1997: АВСТРАЛИЯ.
[8] Limited, T., S. T. C. Limited и P. T. Limited, Scoot Traffic Handbook. (2000).
[9] Мур, Дж.Э. и П. П. Йованис. Статистическое обозначение подрайонов управления движением. Журнал транспортного машиностроения, Vol. 111 No. 3, 1985, pp.208-223.
DOI: 10.1061 / (восхождение) 0733-947x (1985) 111: 3 (208)
[10] Чжан, З.-q., Y. Li и Q. -f. Лю. Изучение техники самоорганизации в системе управления городским движением. Manofacturing Automation, Vol. 2002 г. № 3, 2002 г., стр. 38-40.
[11] Май., Ю. -С. Чиу и X. Ян. Разделение сети управления сигналами городского движения с использованием собственных векторов Лапласа. в 12-й Международной конференции IEEE по интеллектуальным транспортным системам. 2009. Сент-Луис, Миссури, США.
DOI: 10.1109 / itsc.2009.5309560
[12] TRB.Руководство по пропускной способности автомагистралей, 2010 г. Совет по исследованиям в области транспорта национальных академий, Вашингтон, округ Колумбия, (2010 г.).
[13] Саймон, Х.D. Разделение неструктурированных задач для параллельной обработки. Вычислительные системы в технике, Vol. 2 No. 2-3, 1991, pp.135-148.
[14] Фидлер, М.Алгебраическая связность графов. Чехословацкий математический журнал, Vol. 23 No. 2, 1973, pp.298-305.
[15] Фидлер, М.Собственные векторы ациклических матриц. Чехословацкий математический журнал, Vol. 25 No. 4, 1975, pp.607-618.
[16] Фидлер, М.Свойство собственных векторов неотрицательных симметричных матриц и его применение в теории графов. Чехословацкий математический журнал, Vol. 25 No. 4, 1975, pp.619-633.
[17] Вебстер, Ф.V. Настройки светофора. H.M.S.O., Лондон, (1958).
[18] Абдулаал, М.и Л. Дж. Леблан. Модели проектирования сети непрерывного равновесия. Транспортные исследования Часть B: Методологические, Том. 13 No. 1, 1979, pp.19-32.
DOI: 10.1016 / 0191-2615 (79)
-3
[19] Дафермос, С.Равновесие движения и вариационные неравенства. Транспортная наука, Vol. 14 No. 1, 1980, pp.42-54.
[20] Смит, М.J. Взаимодействие на перекрестках и монотонность в распределении трафика. Транспортные исследования Часть B: Методологические, Том. 16 № 1, 1982, стр. 1-3.
DOI: 10.1016 / 0191-2615 (82)
-4
[21] Менегуззер, К.Равновесная модель выбора маршрута с явным учетом влияния пересечений. Транспортные исследования Часть B: Методологические, Том. 29 № 5, 1995, стр. 329-356.
DOI: 10.1016 / 0191-2615 (95) 00011-2
[22] Шеффи, Ю.Городские транспортные сети: анализ равновесия методами математического программирования. Прентис-Холл, Englewood Cliffs, (1985).
[23] Смит, М.Дж. И Т. ван Вурен. Равновесие трафика с отзывчивым управлением трафиком. Транспортная наука, Vol. 27 No. 2, 1993, pp.118-132.
DOI: 10.1287 / trsc.27.2.118
[24] Цзэн, Ю., и другие. Метод распределения городского движения с учетом задержки сигнала на перекрестках. в 10-й Международной конференции китайских специалистов в области транспорта 2010. Пекин, Китай: Американское общество инженеров-строителей.
Произошла ошибка при настройке вашего пользовательского файла cookie
Произошла ошибка при настройке вашего пользовательского файла cookieЭтот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файлах cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
парные благодаря классификации перекрестка.
Этот метод разработан для данных, в которых можно ожидать
множества граничных случаев, участвующих в границе пересечения
. Мы протестировали метод на
примере двух пересекающихся кубоидов, выровненных по краям,
, где каждый сегмент границы пересечения
представляет граничный случай. Мы также протестировали метод
на сцене, состоящей из нескольких Стэнфордских булочек —
nies и Стэнфордских драконов (Levoy et al., 2005) на
демонстрируют правильное поведение метода в более крупной сцене
, содержащей множество границ пересечения.
Наши эксперименты показали, что метод
может найти правильное решение даже в этих нетривиальных ситуациях.
Правильное поведение метода может быть гарантировано только в том случае, если максимальная ошибка вычисления ε на
меньше, чем самый короткий край сетки. Для мод.
els, которые содержат почти вырожденные ребра, алгоритм
может работать некорректно, потому что ошибка вычисления
может испортить выходные данные метода.
Поскольку метод работает с арифметикой одинарной точности с плавающей точкой
точек, он может быть реализован на графическом процессоре
, где операции с более высокой точностью могут стоить очень дорого. Преобразование алгоритма
для возможности его запуска на графическом процессоре является одним из возможных вариантов
для этой работы.
БЛАГОДАРНОСТИ
Работа поддержана проектом SGS-
2016-013 — Advanced Graphical and Computing Sys-
tems.
СПИСОК ЛИТЕРАТУРЫ
и
´
ujar, C., Brunet, P., and Ayala, D. (2002). Топология —
, уменьшающая упрощение поверхности с использованием дискретного твердотельного представления
. ACM Trans. График, 21 (2): 88–105.
Аттене, М. (2010). Легкий подход к ремонту —
оцифрованных полигональных сеток. Визуальный компьютер,
26 (11): 1393–1406.
Аттене М., Кампен М. и Коббельт Л. (2013). Ремонт угольной сетки Poly-
: перспективы применения.ACM
Вычисл. Surv., 45 (2): 15: 1–15: 33.
Бишофф С. и Коббельт Л. (2005). Консервация структуры-
ремонт CAD моделей. Форум компьютерной графики,
24 (3): 527–536.
Бишофф С., Павич Д. и Коббельт Л. (2005). Автоматическое
восстановление полигональных моделей. ACM Trans. График,
24 (4): 1332–1352.
Botsch, M., Pauly, M., Kobbelt, L., Alliez, P., L
´
evy, B.,
Bischoff, S., и R
¨
oossl, C. .(2007). Geometric Mod-
eling на основе многоугольных сеток. Этот документ
поддерживает курс SIGGRAPH 2007.
Брошу Т. и Бридсон Р. (2009). Робастные топологические операции для явных динамических поверхностей. SIAM Journal
по научным вычислениям, 31 (4): 2472–2493.
Кристиансен, А. Н., Нобель-Йоргенсен, М., Оге, Н., Sig-
,mund, О., и Берентцен, Дж. А. (2014). Оптимизация топологии
с использованием явного представления интерфейса.
Struct. Многопрофильный. Оптим., 49 (3): 387–399.
Клаузен, П., Вик, М., Шевчук, Дж. Р., и Обрайен, Дж. Ф.
(2013). Моделирование жидкостей и взаимодействий твердое тело-жидкость с помощью лагранжевых сеток. ACM Trans. График,
32 (2): 17: 1–17: 15.
Edelsbrunner, H. and M
¨
ucke, E. P. (1990). Симуляция
простоты: метод, позволяющий справиться с вырожденными
случаями в геометрических алгоритмах. ACM Trans.График,
9 (1): 66–104.
Энрайт Д., Федкив Р., Ферцигер Дж. И Митчелл И. (2002).
Гибридный метод установки уровня частиц для улучшенного захвата интерфейса. Журнал вычислительной физики,
183 (1): 83–116.
Джу Т. (2004). Надежный ремонт полигональных моделей. ACM
Пер. График, 23 (3): 888–895.
Карасик М., Либер Д. и Накман Л. Р. (1991). Ef
эффективная триангуляция Делоне с использованием рациональной арифметики.
ACM Trans. График, 10 (1): 71–91.
Левой М., Герт Дж., Кёрлесс Б. и Пулл К. (2005).
Стэнфордский репозиторий 3D-сканирования. http: //
graphics.stanford.edu/data/3Dscanrep/.
Ло, С. и Ван, В. (2004). Быстрый надежный алгоритм для
пересечения триангулированных поверхностей. Разработка
с компьютерами, 20 (1): 11–21.
Маклаурин, Д., Маркум, Д., Ремотиг, М., и Блейдс, E.
(2013). Ремонт неструктурированной треугольной сетки между
секциями.Международный журнал численных методов —
ods in Engineering, 93 (3): 266–275.
Мейстер, Г. Х. (1975). У полигонов есть уши. The American
Mathematical Monthly, 82 (6): 648–651.
Мишталь, М. К. и Берентцен, Дж. А. (2012). Топология-
адаптивное слежение за интерфейсом с помощью деформируемого plicial комплекса sim-
. ACM Trans. График, 31 (3): 24: 1–
24:12.
Мурали Т. М. и Фунхузер Т. А. (1997). Создает
твердотельных и граничных изображений палатки на основе произвольных
полигональных данных.В Proceedings of the 1997 Sympo-
sium on Interactive 3D Graphics, I3D ’97, pages 155–
ff., New York, NY, USA. ACM.
Норуддин, Ф. С. и Тюрк, Г. (2003). Упрощение и переделка пары полигональных моделей
объемными методами.
Транзакции IEEE по визуализации и компьютеру
Графика, 9: 191–205.
Ошер, С., Федкив, Р. (2002). Методы набора уровней и динамические неявные поверхности Dy-
(Прикладные математические науки
ences).Springer, издание 2003 г.
Wojtan, C., Th
¨
urey, N., Gross, M., and Turk, G. (2009).
Деформируемые сетки, которые разделяются и сливаются. ACM Trans.
График, 28 (3): 76: 1–76: 10.
Захареску А., Бойер Э. и Хорауд Р. (2011). Topology-
адаптивная деформация сетки для эволюции поверхности, мор-
phing и многоэкранная реконструкция.